ǰҳ
Ŀ¼
��ҳ
�����¸��������ġ�ϸ���ڷ����ۡ�һ��������ȷ�ر��������Ĺ۵㣬���Ŵ����ָ�ɵ�λ��״��ÿ����λ��״�������Ŵ��������ⶩ������ƻ���������ͬʱ��������ѧʵ���о�������ֱ��1892�������ؿ�չ�ӽ����飬��ʼʱ���õ�����ƿ�ݣ�Silene������ڡ��¼��ݵ�ֲ�1894������536��F2����ƿ���з���392����ë��144����ë��2��72��1����1895���������F2�������з��ֻ����кڰߵ�158�꣬�װߵ���43�꣨3��67��1����1896�������ְװ�����Ǵ�һ������������Щ���е���������Ҳ��֤ʵ����Щ���֡�1899����¸�����30�����ϵ����ֺͱ����й۲쵽���Եķ��������������Ϊ����Ӧ��״�ķ������ij��һ��Ĺ��ɲ���Ϊ�г�����ɷ�����Щ�����1900��3�����ڼ�������֮���Ⱥ�д������ƪ���¼����䷢�֣���ƪ���������ѧԺ������1900��3��26�յĻ�������������һƪ�����¹�ֲ��ѧ�ᣨ3��14���յ�������Krizenecky��1965������������������³�������ڣ�4��21��֮ǰ��ʵ���ϱȷ����¹��ij���ʱ�䣨4��25�գ��缸�졣�ڼĸ��¹�����ƪ���������ڽ�ע��д��������������˴����鲢������Щ����д����ƪ���º�ŵ�һ��֪���ϵ¶������¡���Olby��1966��129�����ݴ����ļ��֤�ݶ϶����¸���ܿ�������1896���1897��Ͷ����ϵ¶������¡�zirkle��1968������Ϊ��1899�꣬Kottlet��1979�����ݽ�һ����֤��ָ��ʱ����1899�ꡣ
��������Щ���е¸����ڽ����������Ȼ�������Լ����������ģ�active��A����DZ�ڵģ�latent��L�������������ϵ¶������Ժ����ԣ���ʾ����ͼ�����Է����ò�ͬ�İٷֱȣ�77��5����22��5����75��5����24��5������ʾ���ƺ����������˽���������ԭ��ֵ���ᵽ���ǣ��¸������¼��ݽ��й��������ӽ����飬��1900�����������ֻѡ���ˣ��������¼���X�����¼��ݣ�������ӣ����������¼�������������������ֵ�Ψһ�����Ļ���ͻ�䡣�������ںͱ���ɭ��ͨ��������ȷ˵���ģ�����������״��������״��������ֻ�к�������ϵ¶����ɡ�
�����¸�����˵��������1892������һƪ���µIJο������з����ϵ¶���������¼������Ȼ����1892���Ժ�ļ����вο�����������ƪ���²���ʹ���Ķ��ϵ¶���ԭ�ġ��������ʣ�������ʱ���Ѿ�֪�������ֵ���������ڽ�֮����Ϊ3��1��ֵ���Լ����ԵĴ�һ�����������Ⲣ��һ����ζ����Щ���ֻ��ʹ��������ԭ�ȵĴ���������19����80��������������о���һ�����¸���ԭ����Ϊ��״�������ɶ��������Ƶģ�����ʮ���¡�������394��144��158��34����77��5����22��5%������һЩ��ֵ�����Ÿ������Ӿ�����״������˵�������塣�����ñ�ֵʱ���¸���ָ����2��1��4��1��Kottler��1979���������ϵ¶������£��Ƿ��ʹ��������ԭ�ȵ�ѧ˵�����ܡ�����ÿ��˫��һ�����Ӿ���������״�����ϵ¶�ѧ˵�����ǽ���Զ��֪������Ȼ��ˣ����Ǿͱ�����ܵ¸����˵�������Ǵ����Լ������顰���۳������붨�ɵģ������ϵ¶������Ƶ��������ó�����ѧ˵һ�����¸���ר�������ڵ�λ��״�������Է�������ȷ�dz��ӽ�������Ľ������ǰ��һС�����ܷ�����ԭ��ѧ˵�е����һ�����֣����ӵľ������ƣ���Ȼ������ɭ�ڶ����¸����������ǰ��Ȼ�д������ϵ¶�ʽ�ı�ֵҲû�������ϵ¶������Ľ��͡�
�����¸�����ϵ¶������ȣ���Ȼ�ܾ�ɥ���������������ȥ̽�����ķ��ֵĸ���̵��Ŵ������ת��ӽ����ǶȲ��ͽ���ͻ���һ��ԭ�������γ��ƺ�һ��������Ҫ���ĵ����⡣�¸�����Ȼ��Ϊ�ϵ¶��Ŵ�ֻ�Ƕ����Ŵ�����֮һ����������������ڸ�����ɭ������˵�ġ����ҿ�����Խ��Խ������ǣ��ϵ¶�ѧ˵���ӽ��ձ���ɵ�һ�����⡣����ˣ��������������ϵ¶�ѧ˵��ȥ�о�����Ϊ�Խ�����Ϊ��Ҫ��������ʽ���Ŵ���
�������������ɱ������¸��ォ��Զ���Ŵ�ѧʷ��ֵ�ü����ΰ��ѧ�ߣ�
������1�����������ϵ¶�����˽�����֮��IJ���ָ�ɵ�λ��״�Ĺ۵㣻
������2����������һ��Ⱥ��ʽ������ֲ����֤ʵ���ϵ¶����������Ǵ��ڵģ�
������3������չ���Ŵ���λ��ͻ���Ը��
�������������ֹֻ���ϵ¶��ķ�����֮һ����Ȼ�¸�����ϵ¶���ռ�����ơ��������õ�ʱ��ϸ��ѧ�о��³ɹ�����չ����ѧ˵�����ϵ¶����ǵع���˶��Ŵ������ӡ���Elemente�����ʼ������ʻ�����̽��ʱ���¸���ȴ��֮�����¶����˵Ĵ���ĵķ�����ϵ���������Ŵ�������ԣ��¸����ۺ��˴�������ϵ¶���
�����ϵ¶��Ŵ��ĵڶ�λ���·����߿���˹��Carl Correns��1864��1933��������ͼöࡣ����˵���ϵ¶��ķ���ѧ˵��������1899��10�µ�һ�죩�������ڴ��ϵ�����ʱͻȻ���������Ƶġ����������Ժ�������ʱ��æ�ڱ���о�ֻ���ڼ�������֮��Ŷ����ϵ¶������ģ���1899��12�������������������ָ�������������յ��¸���ķ��������裩��ѧԺ���µĸ�ӡ��ʱ��1900��4��21�գ����ţ���һ��֮�ڣ�������������д�����²��ڵ¹�ֲ��ѧ��4��27�յĻ���������������Լ��5��25�ճ��档����˹��һ��ʼ�Ͳ���Ϊ�������·����ϵ¶���������Ҫ���á�������һ��ͨ���ı����о��õ��ǡ��ϵ¶���������Ϊ�������Լ���˵���·�����Щ�������ѵ������Ͷ����ɸ������߱�������ѧ�Ĵ����о���������κ˹���Ĺ�������֮�ϵ¶��Ǵ������ˡ������ڿ���˹�������·����ϵ¶������⣬Ψһ���Ի��ɵ��ǣ������ڸ����ѧ���������������ڸ������Ů�����������֪���ϵ¶��Ĺ�����Ȼ�����ֿ�����������ʹ�˽��ܵģ��������˹���ϵ¶��Ĺ�������20��ǰ���Ѿ�֪������ȴû�а���һ�������翪չ�о��Dz����Ƿdz���ֵ��¡�
��������λһֱ����Ϊ�Ƕ������·����ϵ¶����ɵ����ǰµ���ֲ�����ּ���Ъ���ˣ�Erich Tschermak��������Stern��1966��ָ��������Ъ������Ϊ���·�������û��ʲô���ɵġ���ȷʵ�����ϵ¶������£���������1900�귢���������б����������˽��ϵ¶��Ŵ��Ļ���ԭ����Ȼ����������ֲ�����ּ�ע���ϵ¶��Ŵ�ѧ����Ҫ������ȴ���˻������á�
����Ϊʲô�ܶ����ڵ��ϵ¶�������ǡ�ɶ���ֲ��ѧ�ң��ϵ¶����¸������˹����Ъ���ˡ�Լ��ѷ������������Ҳû�н��������Ҳ������ֲ�����������ֲ���и�����ѡ�����ֵĴ�ͳ����Ϊֲ��ȶ�����������������֡�Ҷ�������ܱ���ţ�����ȼ�����и���IJ�������״���������ּ����о��Ĵ������״���Ǹ߶ȶ�����Եģ����������ڽ��м��ϵ¶�ʽ������Ȼ����1900���������ɭ��ʼ�о����ݣ�Cuenot�ڷ����Ϳ�������19o2����������ʼ�о������࣬1905�꿭�������ù�Ӭ��Ϊʵ�鶯������Ŵ�ѧ���о��ܿ������ֲ���Ŵ�ѧ������Ħ��ѧ�ɺ�����ά��ɷ�ѧ�ɿ�չ�����ǵ��о�������ͳ�����ֲ���Ŵ�ѧ������1914�꣬A��Lang��������1900���Ժ��鶯���Ŵ�ѧ���о��ɹ���ר�۾�Ȼ����890ҳ��ƪ����
����ֲ������ߵ�ֲ��Ŵ�ϵͳ�Ķ�����ԶԶ���ڶ�����Ҫ�����ձ���ɵ�����˵�����ײ�������������ɽ��������Hieracium����������ֳ�����ں���ֳ��apomixis��ϵͳ��ʹ�ϵ¶����о�����ۣ��¼��ݵľ����Ӻ�Ⱦɫ�廷���µ¸�������˴���������γ�ѧ˵���Ի��ܷ۵Ľ��ڴ��ϵIJ˶���Phaseolus��ʹԼ��ѷ�������Ȼѡ������á�ϸ����Ӱ����ֲ������Ȼ�ȶ����и��ձ��������ֲ���Ŵ�ѧ�ң��ر��ڵ¹�����ע������ȫ�������ⷽ�����£��ڷ����Ŵ�ѧ��ǰ��û�������ر���Ҫ���о��ɹ�����һ���棬ֲ��粻���ṩ���㶹���һ��й����ࣨ�ر���С�������ף��������̲��Լ��������������Ŵ�ѧ�����о���ֵ�����֡�Ŀǰ��û���˴������Ŵ�ѧ�о��������õĸ��ֶ���ֲ����������������ıȽ��о���������ϣ����������ֻ��ӡ֤�Ѿ��ɹ�Ӭ��������ȷ������ʵ���ڷ����Ŵ�ѧ��ǰ�����Ŵ�ѧ�о�������ֲ��ѧϵ����ѧϵ�ֱ���У�ֲ���Ŵ�ѧ�ߺͶ����Ŵ�ѧ��֮���ѧ������������������ϣ����������Ծ��30����Ժ�͵�ֲ����ࡢ�������ĸ����ԭ�����ϸ���������������Ϊ�Ŵ�ѧ�������ӵ�ʵ����ϡ�������ʶ�����������ԭ���������Ŵ�ϵͳ�ϵ��ش���60����Ժ����ٶ�ȼ�����������Ŵ�ѧ�о�����Ȥ��
�����Ŵ�ѧ��������ʷ���Է�Ϊ�����Σ�ͷһ�δ�1900�굽1909�����ң��ڶ�����ʼ��1910�ꡣͷһ����������Ϊ�ϵ¶�����ʱ�ڣ���Ҫ�����������йؽ���������������Լ��ϵ¶��Ŵ��Ƿ��ձ���Ч�����⡣��һʱ�ڣ��Σ�����Ҫ���������ǵ¸������ɭ��Լ��ѷ����������Ϊ�����ڵ��ϵ¶������ߡ�����ͬ���˶ԡ��ϵ¶����塱����в�ͬ�����⣬������Ҫǿ�������ϵ¶�������ĸ�����������Խ����Ŵ�ѧ��ѧ�߶��ԣ���ָ���ǿ����Ŵ�ȷ�������ѳɶ�����������Ӳʽ�Ŵ���ʱ�ڡ����ڽ�����������˵�����ϵ¶����塱ָ��������һ��ʱ�ڣ�����һЭ����ijЩ�������Ŵ�ѧ�Ҵ����Ź��ڽ�������������γɵ���ȫ����Ĺ۵㣬����һʱ����ͻ��ѹ������ΪԶ����Ȼѡ����Ҫ������Щ�۵�Բ���ѧ�����Ǹ����ġ���ˣ����ϵ¶����塱��ͬһ������ʱ��������ʾ������֧�֣���ʱȴ���б��塣
�����ڶ����δ�1910�꿪ʼ����Ҫ������Ħ��ѧ�ɣ���Ҫ�����ڴ����Ŵ�ѧ������о����������ı��ʡ�������Ⱦɫ���ϵ�һ���еȵȡ��ɱ���ɭ��1906�꽨��ġ��Ŵ�ѧ����genetics������ʺ������ձ���ܲ���Ϊ�о��Ŵ��������ſ�ѧ�Ĺ�����
�����ϵ¶���������������Ȼ������34������·��֣���һ�����·���֮��ȴ��ǰ��δ�е��ٶȹ㷺����������˹����Ъ���˶�����1900��4��δ�����¸�������¡����ֱ���5�º�6�·������Լ����й����¡�����ɭ��5��8����Ӣ���ʼ���ѧ��Ļ�����ͨ�����ϵ¶������飬�ڷ���CuenotҲ�ܿ�ͽ������ϵ¶��Ĺ�����
������������Ҫ�Ŀ�ѧ���£����Ľ�չ��ͷ�ڲ�ͬ�Ĺ���Ҳ������ͬ���������ʣ����ϵ¶��Ŵ�ѧ�Ľ�չ����Ӣ����ңң���ȵģ������ü���������������������Ĵ���������Castle��East��Ħ���Լ�����ѧ�ߣ����¹����Ŵ�ѧ��Ȼ�̳���19����8O����Ĵ�ͳ�����ط����Ŵ�ѧ��һЩ���������Ŵ����������Ļ�����ϵ�ϸ�����Ŵ���ԭ�������Ŵ��ȵȣ����ڷ�����Cuenot����һ����ͷ֮��һֱ��20����30�����û�ж����Ϊ��������������Gaissinovich��1971��ָ���ģ���ֻ�ǵ�����ά��ʱ���Ŵ�ѧ�ŷ�չ��Ϊһ�ſ�ѧ����������ŷ��û���Ŵ���ѧ�������Ŵ�ѧ��ʲô�ط��������ٺͰ��ĸ�����չ����ȫȡ������һ����Ĵ�ͷ�ˡ�Ȼ����ֵ��ǣ�����˹�͵¸����������ϵ¶��Ŵ�ѧ��չ�϶�û�з�����Ҫ���á���һ���������������ڣ�������ڱ���ɭ��1861��1926���������ϵ¶��Ŵ�ѧ��Ҫ�������ʶ�̶�Զ����ν�����·�����֮�ϡ�
��������ɭ�Դ��ڻ��ս�˹��ѧW��K��Brooks���ڵ��о��Ҷ����ڼ䣨1883��1884���ͶԲ��������죨���ڶ��ࣩ������Ȥ����80�����ͽ����������飬���������о��������ⷽ����о�����1897�����ҡ�1899��7��11������ʼ���ѧ��������һƪ��Ϊ����Ϊһ�ֿ�ѧ�о��������ӽ����ӽ����֡������ġ�����ƪ���Ŀ��Կ�����ʱ����û������Ŵ�ѧ˵���������������������ϵ¶��۵�������͡�ֱ��1900��5��8�����ڴӽ��ŵ��صĻ��϶����ϵ¶���ԭ������������ϡ����ܿ�ͳ�Ϊһλ�ȳϵ��ϵ¶������߲����ϵ¶������·���������Ͻ�ע�����ڻʼ���ѧ����־�ϣ�1900�꣩������ɭ������һ���ֳ�������Ϊ�ϵ¶�����ѧ˵�Ƕ����ģ�����ģ��۵㡪���������γ��Dz���������Ľ���������Ŀ϶����¸���Ҳ��������ƵĽ���ѧ˵����Ҳ��Ϊ�ϵ¶��Ŵ����ӵIJ���������������������γ�ѧ˵����Ҫ֤�ݡ���ˣ��ϵ¶�ѧ˵֮��������㷺ע�⾹Ȼ���������Ƕ��ǵر���ģ�������Ǵ���ģ����⡣���ڱ���ɭ��¸�����۵�������ķ������ڵ�ʮ���¡����ܣ���������ֻ���۱���ɭ�Դ����Ŵ�ѧ�����Ĺ��ס�
�����Ŵ�ѧ�еĴ������Ҫ���ﶼ�DZ���ɭ����ġ���Ϊ������ѧ�ƴ����ˡ��Ŵ�ѧ������´ʣ�����1901���״��ˡ���λ����allele��ԭΪallelomorPh��������������ӡ������Ӻ��ӡ���������Щ������ȷ��������ٽ������ʱ�ڵ�ѧ����������Ȼ������ɭ����ͬ�»��������˽��Ŵ�����������ʵ���ԵĹ��ס��������ȷ������ϵ¶����۲쵽�ļ�����������������������ȫ���������Ŵ�ѧ����ͨ������ɭ����Ӣ��ȡ������ŷ��������������ȫû�е���ͷ������
��������ɭ��һ���ܸ��ӵ�����ڱ����кö��������ֱ�����ͬʱ������ȫ��������ҵ�����Ǽ汣���������һ������ֻ�����1900���Ժ��ͷ10���У������Ŵ�ѧ����Ҫ��ң�Caslle��1951����˵��ʵ���Ǻ��е����ģ���˵����ɭ�����Ŵ�ѧ��������ʼ�ˡ���Ȼ��1910���Ժ�������Ⱦɫ��ѧ˵�Լ��������������Ȼ�γɵĹ۵�Ͳ�����˵�ǽ����Եġ���Ϊһ����������˵�����������˲�������ԣ�1908��22����������ϧ�������ֵ�������������û�����⣬�����ͻ�����˿�����ζ˭Ҳ��Ը��ȥ��һ���ƶ���ǰ��������Щ���������Զ��¶����Ŀ��֮�¡�����������еĴ��õĴ���ʯ�ϣ�����������������һ���ӹ���ָ����һ������Ӧ��������ʲô�ط����������Լ����о������У����ͷdz�ע��ʵ���ϵ����������ϵ����⣬����һЩ��Ҫ���־�����ѭ��һ���ԵĽ����
�����ϵ¶��Ŵ�ѧ�Ľ�չ
����1900���Ժ��Ŵ�ѧ���·��ֳ��ֵ��ٶ��ڿ�ѧʷ�ϼ�����û�������ġ������Dz���Lock��д���Ŵ�ѧ�̿��飨1906�����������е�163��275ҳ�����DZ���ɭ�Ľ̿��飨1909�������Ǿͻ�Ϊ1900������Ƕ��ϵ¶��Ŵ�ѧ����ij���̶ȶ��е����ȡ��������Ѹ�ٵ�ԭ����ʲô��ԭ��֮һ��Ȼ������ѧ˵�����������ͼ����������κ���ȥ�����Ŵ���������֤���Ƿ��ձ���Ч����������һƬ�¿��ص��������κ��˶���ȡ���·��ֵĻ��ᡣ�ϵ¶����ɿ��Զ��Ŵ���ʽ����Ԥ�Ⲣ����ЩԤ���������Լ��顣��һ��ԭ����û�ж��ۣ�����Ϊ1900����ǰϸ��ѧ�о���35������ȡ�õĻԻͳɾ͵춨���ι̻���Ӧ���ܹ���ϸ��ѧ���ر��Ǵ�Ⱦɫ��ĽǶ���˵������һ�д������Ե��Ŵ�ѧ���֡�Ⱦɫ��ϸ��ѧ�ѳ�Ϊͨ������ѧ����������������������������ܹ�ʹ��֮ǰ���ѽ��ɡ�Ȼ����ֵ��ǣ���ʹ������ʹ��֮��Ҳ�������Ŵ�ѧ�ҡ�����Ħ��֮ǰ�ı���ɭ����������East����ȫ�����ˡ�
�����Ŵ������йػ��Ƶ�֪ʶ��Ӧ��������ѧ�ĸ�����ͬ�������������ѧ������ʮ���¡���ʮ���¡�������������ѧ�����潫�������۴����Ŵ�ѧ���档
���������ԣ�Semidomhance��
�������ϵ¶����������߶���״�У�����ÿһ����״ֻ�ֱ�����������壬�����Եĺ����Եı����塣���������ϵ¶����������ֵ����������������е���״�ԣ��ɶ���״��������ˡ�������̸�����ھ͡�����ǡ�ô����ױ�ֲ��֮�䡣������˹��1900��Ҳͬ������ijЩ���Ӳ�������ȫ���Զ��ǡ������ԡ������������F1�����Ͷ��ٽ���˫��֮�䡣����֮����ɭ���ðҼ��ͺڼҼ��ӽ�ʱ������������ɫ�İ���·���ǣ����������Ҽ������İ����ԡ�
�����ⲻ��֤ʵ�˰������Ǵ��ڵĶ��һ������ϵ¶����ɶԶ����ֲ�ﶼ���á�������ͬһʱ��Cuenot���ݶԼ���ëƤ��ɫ������о�Ҳ��֤����һ�㡣���ڶ����ֲ���ϸ����ϸ������ʾ��ȫ��ͬ��������һ��ʵ����һ���ֿ��ܲ�����ȫ�������ϡ�Ȼ���ϵ¶����Ŵ�����ͬ�������ڶ�����ֲ������һ���ֽ�һ���ݻ��˴����ڶ���ѧ��ֲ��ѧ֮��Ĺ��Ͻ��ޡ�
���������Ŵ���λ
������1909����ǰ������Ϊ�ɼ���״�������Ŵ����ӻ�û��һ�����ϵ����˹���������˶�������ġ��¸��κ˹���Լ����������Ŵ������ѧ�߶��ٶ�������ij�־��в�ͬ���ʵĿ������ʣ�����������ʹ�õ����Ʋ�δ���㷺���ã�����ʮ���¡�����
�����ϵ¶��������о������н����Ŵ����ʱ��ʵ������ϸ�����������ȣ�����1865�굱ʱ��ϸ���˺�Ⱦɫ����˽�������ޣ��������������Ƿdz����ǵġ�������������ָ����������MerKmale��������״����Charaktere�������������ڱ����Ͳ�Σ���Ȼ�����õķ���A��Aa��a���ձ���Ϊ��ָ�Ŵ��͵Ľṹ���������ĵĽ����У�1866��41��42������ʹ�á����ӡ���Elemente����ʴ�10��֮�࣬�����м��κ���������ʹ�õġ�������ʵĺ���ʮ�����ƣ��������Ŵ����ʲ�û�������ĸ�������ϵ¶�������������ģ������ڵ��ϵ¶������߿������������ľ�������������˵���ϵ¶��Ŵ���
������Ȼκ˹��������ʾ������������֮������𣬵���ֱ��190O�껹û�С������͡��͡��Ŵ��͡�������ѧ�����ʡ��͵¸�����Ŵ����ʺ����壨�����ͣ���û��ʵ���ϵIJ��죬��Ϊ��������ķ��ӿ������ɵش�ϸ��������ϸ���ʡ�����Ϊ���Ӿ��ǵ�λ��״�������״�������Ŷ�ÿһ�������Ŵ�����״����һ���������Ŵ��������¸�����ʱҲ���Ŵ�Ҫ�أ�genetic elements�����������ӡ���factors��������ɭ��Ħ��ѧ�����Ҳ��������һ���ơ�
�����͵¸�����£�����ɭҲ�ֲ�����Ϊ�������Ŵ���������Ϊ����ı�������״֮���������������λ��״�������ǡ����ӽṹ�ж�����һ�Ǵ˼��˵ģ���״������1902����Ϊ���ܹ�ָ�������Ķ�����һ����������㶹�е�Բ�����嶹������ɭ�����ˣ�ϣ�����б�ʾ�˴˻�����ֲ�����ת������������ʡ���all��elomorph�����Ϊallele������Ϊ��λ����Ȼ�����ֲ����������壨���ʣ���״�����������еĶ��ӣ�������������ԭ����1910��֮ǰ�����ձ�ش����һ��Ĭ���������Ŵ����ӣ���������״֮�������1��1�Ĺ�ϵ����˵��ᵽij����λ��״ʱ������ָ�Ŵ���������������ͱ��ֶ��ؽ�Ҫ���������������Զ���Ĭ����ʹ��������������ġ���Ⱦѧ˵����contsmination theory����
��������1900���Ժ��Ŵ�ѧ���Ѹ�����࣬���б�ҪΪ���Զ����Ŵ�����״�����ʻ����ƶ�һ����������Ŵ�ѧ��Լ��ѷ��W.L.Johannsen��1857��1927�������ϵ¶�����������������¸���������ķ��Ӻ����ƣ������1909�꽨�齫���ӣ�pangen������ּ�Ϊgene��ʾ�Ŵ���״�����ʻ�����Լ��ѷ��һ�����������ߣ�������������ҪΪ��������ʵĶ���Ϳ���ȳ���ɫ�ʡ���ָ�𡰽�������Ϊ�����Եġ�����̬�ϱ�ʾ�ṹ�����ĸ���������Ŵ�ѧ���Ȳ���չ�dz��к��������������������1909:375�����������û��Ϊ�����¶����ֻ��˵���������������Ϊ������λ��Rechnungseinheit����������Ȩ������ĵ�ѿ��κ˹������Դ����ӻ����������Ƶ������Ը��������Ϊij����̬�ṹ������Ҳ��Ȩȥ����ÿһ����������൱���ض��ı�������״������̬ѧ������˵�ģ��������������������1909����
����������巴ӳ���Ǹ�ʱ�ڹᴩ������ѧ�����е�������硣���������ߣ�Ҳ����Լ��ѷ�����������ܽ����Ĺ�ϵҲ�������ǵ�Ӱ�죩Ҫ����ѧ�۵����һ�С���̥ѧ�߳��ں����ۣ������ۣ���ͳҲͬ����Ը����ܿ����Ի�����Ϊ��ʹ�����������ȳ��ۡ�Ħ�������Ը����ϻ����������ǿ����Ի���Ҳ�������������Ŀ��ǡ�����б����۵�ijЩӰ�죬�������Ƿ��Խ����ֵı��ʼ��Էָ�ġ�1917��Goldschmidt�����������Ŵ�ѧ�ҶԻ�����ֽ�С����̬�ȣ���������Ϊ�Դ��������������̬��������Լ��ѷ�Ի����ʲ�ȡ����֪�۵Ľ���������Ͳ�����ij�����صij羴���飬�Ի�����������ԵĹ۵��ʾ���ʹ��������Ȼ�����������ʵ���ʱ�����֤��ǡ�þ���Լ��ѷ�dz���ϸ�ش����Ķ������ų�����Щ���ṹ��������ʵ���ϣ���Ħ����ʼ�������յ���ɭ�Ϳ����һֱԽ��Խ�ӽ�����Ľṹ���Լ��ѷ���õġ������������ܿ�ͱ��ձ���ã���Ϊ�������˶Ա�ʾ�Ŵ���λ�������������Ҫ��Ȼ������ȱ���ϸ�Ļ����壬��ͳ�Ϊ��������ijЩ���۵IJ���ԭ����˼����ҵ���һ��ԭ���ǣ�����һֱ������ѧ���ǶԻ���ĺ�����ʲô���Ҳ��һ�¡����統̸����Ӭ�İ��ۻ���ʱ���е�ѧ����Ϊָ���ǰ��۵�λ������һЩ������Ϊ��ָ����ͻ�䷢���Ļ�������λ�㣩��Ҳ�������а��۵�λ����Ļ�������
����Ϊ���ɼ��ģ������ṹ���Ŵ���λ���조�������ֿ�ʼ��ֱ������˽����ı��ʣ���ε�·�������ģ�Ҳ�Ǽ��ġ��������Ŵ�ѧ�ң�����������ǰ�е������ա���H��J��Muller��ʵ�����ǰ����ǵı�����ѧ��ҵ��������һ̽���ϡ������н�����������֣�50�������Ϊ�������õĸ߷���ȷʵ����Լ��ѷ���ܾ����ϵ����ֽṹ�ϵĸ����Ժ������ԡ�����ȥ���ֻ�������ܿ�ʼ��ȷ��һ���dz���ĥ�˵����⡣Ħ������ͬ��ʮ����ȷ�ؾ������о��ı��˵Ļ�����ͻ�䡱���֣�������Ϊ�������һ����ϣ����Ƕ��Ш�ӡ�
�����������·������ϵ¶����붨�ɣ��Ŵ��Ա������Դ����ͱ�ú�ͻ������λ����Ĵ���Ҫ��������˵��������ļٶ����첻�ϱ������Ա���Ȼѡ���г�ֵ�ѡ�������Է������á�Ȼ����˵�����������Դ����ʱ�ѵ��˽����ĵ�����յ�ʱ�����ϵ¶����������о��������ʱ�����չ������ʵ�������DZ���˷������ϰ���
������Ҫ�ϰ��ǵ�ʱ�о�����Ĵ����ѧ����Ȼ��Ϊ��������졣�������ķ����С��ܶ�ϸ������Գ�֮Ϊ������족����������Դ������������Ϊ������죬�������죬������������졣��������������Ҫ�Ե���ʶ�����Ľ���ѧ˵��֧��֮һ��Ȼ��������ֳ��ϡ���Щ���졭��������ͻȻ�����ġ���һ�������ġ���30ҳ�����оٳ�ת�湷��turnspit dos���Ͱ�������Ϊ����ͻ������ӡ�����ɭ������ͻ���Ϊ���������졣���������������������ƾõ���ʷ���Ͱ���ͼ�ı��ʸ��������й�ϵ�����ʳ���С���ȵ�żȻ�Ա��죬���κ��ش��ƫ��ֻ��ͨ���±��ʣ��������ͻ���ģʽ����ͻȻ��������ʱ��Ϊ����������������ȫ��ͬ���ڽ����е�����Ҳʮ�ֲ�ͬ���ر���������ͳ��ѧ�Һ��ϵ¶�ѧ��֮������ۣ�����ʮ���¡������������۵Ľ��㣬����������ʵ���Ǵ�������ʱ��һֱ��40��������ۺ�ʱһֱ���ϵؽ��С��¸���Ĺ��ڱ����������1909����ӳ�����������������������̶̳ȡ�������Mayr and Prvoine��1980����
��������������������
���������������ʽ�Ŵ��Ͳ��ѽ�������졣�ڲ�������Ӱ�죨��Ӫ���������κα仯����Ӱ��������״��ʹ֮�����ı䡣��������������˵�����������������仯ʹ����Ľṹ�����ı������£������û��õIJ�λ�ֱ���ǿ��ͻ����������ԭ������ṹ�����˸ı�IJ�λ����������ѿ����Ҳ�ᷢ���仯�������dz�ַ��ܺ�ͻᷢչ��Ϊ�µġ��ı��˵Ľṹ����������ʽ�Ŵ�������ѧ��Ҳ��ȡ�����ƵĽ��͡�����״���ּ�������״������Щ�ּ���״֮��IJ���С������Ϊ�������졣����µ��Ŵ��Ա���������ij��δ֪���̣�����ͬ��������ʽ�Ŵ����ּ���ԭ���Ѵ��ڵı��졣����һ���ͳ��������ֵı��ʾ��в����������������������ڽ�����Ҳ�Ͳ������⡣����ֲ�����ּ��ձ���л�����Ӱ���Ŵ������ԵĹ۵㣨Pritchard��1813��Roberts��1929����
����1883�굱κ˹����������ʽ�Ŵ�������ͷ����˸����仯��������������������������µı�������������߱����ԣ���ĩ��������ԭ����ʲô��κ˹���͵¸���Դ˶�û��������ɳ�ֵ�ѧ˵�����ڵ��ϵ¶������ߵ�ʱ������ע�����ڲ��������������ϣ����ٻ���������Ӹ�����졣����ȥ���ϲ��������ϵ¶����Ӻ���������֮���ì����ʹ�������Խ���¡�
����������������ȱ����ʽ�������谭���������Ľ��������ĬĬ�ؽ���������������Ҳ����Ҫԭ����Щ�������������������⣬��������ʽ�Ŵ���������κ˹�����ԣ����ں��Ŵ�����Ȼ�����ϵ¶����ɣ���ģʽ˼���Լ����Ŵ��ͺͱ����ͻ������塣�������������Ѻʹ������ڵ�ʱ��������ֱ�����ֽ������������±�����Դ���Ŵ�ѧ���⡣����Ľ��ʵ����ͨ���о�������������һ�ػط�ʽ����Ȼ���ַ�ʽ��ǰ������Ϊ���������������������ȫ�ء�
��������������
�����Ŵ��˾�����֪����һ��Ⱥ�壨��Ⱥ����ż�����и���������ڲ�ͬ������������Ⱥ�����������������Ұ��������������ֲ������������ж������֡�������Ⱥ����������κα����嶼�Dz�������������ӡ��������ѧ�ж���������ɫ�����������壬��ָ���Լ����ֻ��Ρ�15���ͺ�16�������ڣ�����Ȼ�类�����ġ���ֳ���������������������������ʱ�����ֹ��ﶼ������ֵس����ڸ���ͼ���ϣ����д�����������Ļ��ζ������ͷţ�������������Ǵ��������������ʨ������֮���Ƕ���塣
����159O�꺣�±���ҩ��ʦSprenger������ҩ���з���һ��Ҷ����״��ȫ��ͬ�����°����ˣ�Chelidonium majus��������֮����������Ӽ�������������һ��ʱ�����ŷ������Ҫ��ֲ��걾�Ҷ������ı걾��17���͵Ĵ����ֲ���鼮��Ҳ�ж��������������µı�����һ�㱻�����ǰ���������һ�����֡�310���Ժ��¼�������һ���쳣ֲ����ʾ�˵¸��ʹ�������һ���µ���Ҫ����ѧ˵��
����������ֲ�������Ե��쳣������Ƚϳ�����ʵ���������������ձ��֣��������ڻ�����ɫ����״�ϣ���������Щ����������ġ��ڼ�����ͬ��Ҳ���ֹ���ţȺ�е��Ǹ��壬���Զ���Ϊ������������һ�Ⱥܳ����İ�����ancon��������Ϊ���ر�̲�����Խ��ʻ�ǽ����������������Щ�����У���������ͨ�����״��ؽ���֮�Խ�����ķ���������������ϵ���������ǿ��Գ�֮Ϊ�������ϵ¶��Ŵ����Ϳ˶�·���������ӽ��еķ����෴����û�С��ںϡ�����Ҳ���ظ����״�ģʽ����ֵ��ǣ���ղ��ʹ����֮���������ں��Ŵ������У����Ƕ���ȫ��������һ��ʵ������ʮһ�¡�����
�������������쳣���������������ν�ķ������뻨��Peloria����1741�����ε�һ��ѧ�����������ɳ��������һ��ֲ����Ʒ��������ֲ������ۣ�������ζ���������ࡢ���������ӵ�������ɫ��էһ������ͨ�ĵ��Ʋݣ�Linaria����ȫ��ͬ��Ȼ����ͨ���Ƹ���кͽ�������Ƶĵ��Ͳ��Գƻ������������뻨���Ǿ���5��ͻ��ķ���Գƻ������εó��Ľ����ǡ�������ֲ���ɱ������ӷ�ֳ�������һ�������֣�������һ��ʼ���еġ����������ߣ������εķ�������������뻨������һ�����ֻ����������һ���һ����ȫ�µĸ١��ⲻ����ҡ�����ε����̶ֹ�����ĸ�����һ��ƺ��������ķ���ԭ��Larson��1971��99��104�������������Ϊ����ӽ��йأ������ܿ�ͷ��������ֹ۵㡣���֤���������뻨������ԭ������Ϊ�������ǹ̶�����ģ����κ���������������������ġ����֡������������ġ�ֲ����־����1755����Ҳû���ᵽ����
��������֮���һ�ٶ����������쳣������±��ַ��ֵ�Խ��Խ����ԽƵ�������Ⲣû���ṩʲô�¼�ʶ����������һʱ�������ص�ȴ��������仯���ͱ��輰��ͬʱ������˵��������һЩ������ֻ�Ǻ����ָ����йأ��������Ž���˼����չ�����ּ�����Դ��ʽ��������µ����塣����ǰ��ָ�����¸���������Ĺ�עΪ�ϵ¶����������˴ٽ����á���������Դ�������Ժ�����������Խ��Խ�ͽ����йء�
����������һ���Դ����ԭ��ּ��������˵��һ������������ֵ�ͻȻ������ȫ��һ���鷳��������Щ�����ڵ�������в��Ϸ���������������µĴ������ֲ��հ�����˵���ֵ�ÿ�ο���¡��ڴ�����Ժ�ʱ�ھ���Щ�������DZ������ߵĽ�������������Ҳ��������������Ϊ�����ɴ˿��������γɿ�������Ȼ������Դ���̣�����ʮ���¡�����
����������ر�ǿ�������Ľ����ԣ�Ҳ����˵�����������ڽ����ϵ���Ҫ�ԣ����Ⲣû��ʹ��������ͬʱ�����ŷ��������裬�߶��أ�Kolliker�������ˣ�ƫ��ͨ����������������ʽ�����ֺ�ģʽ��Դ��Ȼ����Ҳû�б���˱ȱ���ɭ�����������ʶ�������������Ҫ���壬�������ռ��˴���������֤�������۵㣨1894��������ʮ���¡�����
�����¸����ͻ��
����һֱ�����·����ϵ¶������Ժ���������Ĺ۵�ų�ַ�չ��Ϊһ����Ҫ�Ľ���ѧ˵�����¸���ġ�ͻ��ѧ˵����1901��1903����ѧ˵�ڽ�������ѧ�е����ü���ʮ���¡������¸������������չ�������Ŵ�ѧ˵ʱ����������չ������ֲ����ֵ��ӽ�����Ҳ�о�����Ȼ��Ⱥ�еı��졣1886�������������¼��ݣ�ҹ���㣩�Ĵ���Ⱥ�������ں���һƬ���ߵ�����������з���������ֲ�꣬����Ϊ������ֲ���������������弫����ͬ���Կ������²��������֡������ڵ¸���������о����Ի��ڷ���Ȼ�����ȶ�����������������ֲ���������������¼��ݵĸ�����Ҳ������������ģʽ�������������������Ҫ�ı��������¸��ﻹ������20�����ϵĸ��������Ϊ�����֣����Ի��ڷۺ�ȷʵ�ȶ����䡣
�����¸���Ϊ�����¡����֡������Ĺ��������ˡ�ͻ�䡱����ʡ����ǵ�������Ŵ�ѧ˵��ʮ����Ҫ���������˵���䡣��ͻ�䡱���������17������Ҷ�ͱ�������ʾ����ľ��ұ仯��Mayr��1963��168������һ��ʼ���ͼ����ڲ��������������ڻ�ʯ�ı仯��1867����ʱ�Waagen��ʽ�����ڹ�����ѧ��ָ��ϵϵ���п��Էֱ����С�仯���ԡ��¸�����˽������÷���Ϊ�����ص��ᵽ����1901��37��Waagen���������ǵ������кܶ�ʣ����硰��Ӧ����һ������ͻ�䡱��ʼ��øɹ������øɹ��̵Ľ�������DZ�������Ӹ����������ǡ�ͻ�䡱��ʱ����ָ�Ŵ��͵ı仯����ʱȴָ�����ͱ仯��������ǣ��ڵ¸�����Ŀ�У�ͻ����һ�ֽ�����������Ժ���Ŵ�ѧʷ����Խ��Խ��Ϊר�ŵ��Ŵ�������ͻ���������ֻ���������������˽���ܶ���Ϊʲôͻ���ڽ����е�����һֱ�������۲��ݡ�
������Ȼ�¸����á�ͻ�䡱����ʱ������ֵ�ͻȻ���֣���������Ȼ���˽�����仯���������ʣ�������ʵ�ϣ����ǽ�֮������ʾ�����͵�ͻȻ�仯�����Ѿ��������о��¼��ݵ�ѧ����֤ʵ������֤���˵¸�����˵��ͻ�伸��ȫ����Ⱦɫ�����ţ�������ԣ��ı��֣����к������ִ���ָ�Ļ���ͻ�䣨�����ģ���
����������ʮ����Ŵ�ѧ�о���ʹ��ͻ�䡱��ʰ�������ԭ���ĺ��岻�壬�͵¸��������Եģ����Dz������ֵĹ��̵�����¸�����ȷ�Ľ�������������ڲ���������ĵ�λ�ϣ���ͻ�䡭�������˱����Կ�ѧ��һ���ر��֧��ͻ��������ɼ��з������Ҽ�Ϊ�������������ı������������IJ���һֱ���֡�������ٶ�������������ɱ˴˽�Ȼ��ͬ��һ����Ŀ�ĵ�λ��ɣ���������Ҫ��֧������ı����ԺͿ�ͻ���ԣ�֮��IJ��ͺ����ԡ�һ���µ�λ�ij��־ͱ�־��һ��ͻ�䣻Ȼ�����µ�λ�ı��ﰴ�����ֵ�����ԭ�Ѵ��ڵ��Ŵ����ӵ���ͬ����Ҳ�ǿɱ�ġ���1901��iv--v��
������Ȼ�¸��������ͻ�������Ľ��������ϵĽ����Ǵ���ģ�������������ǰ���κ��ˣ�����ǿ�����Ŵ���״��������Դ������һ������Ӧ�鹦�������������ϵ¶��Լ��о��Ŵ����������ѧ�߾�һֱ̽��ԭ�Ѵ��ڵ��Ŵ����Ӻ���״�Ĵ��ݡ��¸����ʹ����ע���Ŵ������������Դ���⡣���ܡ�ͻ�䡱����ʵĺ����1901�����������˶�ô��ı仯������ʱ��ͻ��һֱ���Ŵ�ѧ��һ����Ҫ���⡣
�����¸������������Ƕ�ô�ڷܿ̿��Ѱ��һ�������ֲ������ȷ��֤ͨ��ͻ���������Ȼ�γɡ����о���һ�ٶ���ֲ����dz���һ���������������ȫ����������Ϊ���ǵı��춼����������Ԥ�ڵ�����������ȥ������ǿ��ָ���¼����Ƕ�ô���⣬Ȼ������Ȼ����û����ʶ����һ����ѧ˵����ڴӵ��������������۲쵽���������Ƕ�ôΣ�ա�
��������Renner��Cleland��S��Emerson�Լ������Ŵ�ѧ�ߵij�ɫ�о�����֤��������Cleland��1972�����¼�����һ���������λȾɫ��ϵͳ����ϵͳ���ڴ����ӵ���������������Ӻ���������Զƽ��ġ��¸������ͻ�������ʵ��������Ⱦɫ�廷�ķ������������������ֲ�����ֺͶ����У�����ijЩ�����ġ�����ͬ����ƽ��ϵͳ�����⣩�������ڡ��¸����ͻ��Ȳ��������������ԴҲ���������γɵ��������̡�Ȼ�����ġ�ͻ�䡱��һ����ȴ���Ŵ�ѧ�б�����������������ΪĦ����������������Ħ���ǽ�֮ת����ʮ�ֲ�ͬ���Ŵ�����
����1910�����Ŵ�ѧʷ�ϼ�����1900��ͬ����Ҫ������һ��Ħ�����������о���Ӭ�ĵ�һƪ���ġ����·����ϵ¶����ͷһ��10�건��ɭ���Ŵ�ѧ�ķ�չӰ�켫����������ͬ�²��������֤���ϵ¶����ɣ����һ����ֺͽ������������������������⡣����ɭ���Ŵ�ѧ�Ĵʻ㷽��Ҳ��������Ҫ���ס�����10���в�������Boveri��Ҳ֤ʵ��Ⱦɫ��������ԺͶ����ԣ������ԣ�individuality�������ܻ�ӭ��
������̥ѧ��Ħ����ȫȻ������ɪ��-��������Sutton��Boveri��Ⱦɫ��ѧ˵��ѧ��֮һ����������ѷ��E��B��Wilson������ŦԼ���ױ��Ǵ�ѧ��ͬ�¡����DZ˴�֮����Ȼ�������Ȼ����ʱ���˶�Ⱦɫ�����Ŵ�֮��Ĺ�ϵ�Ľ���ȴ��ȫ��ͬ��1908��Ħ����ʼ�����Ŵ�ѧʵ�飬����õ�ʵ�鶯���Ǵ����С����������о�������ľ��������Ƿ������ò����ද����ʵ�飬��Ϊ���ǵ�����ʱ�䳤���������øߣ�����������������ʱ��������λ�����Ŵ�ѧ�ң�W��E����������Frank Lutz���Ѿ�������ͨ��Ӭ��Drosophila melanoggaster������ʵ����ꣻ���ֹ�Ӭÿ�����������ھͷ�ֳһ�������ӵ��ķ�ţ��ƿ�Ϳ����������Ҽ�����ȫ���ܲ�����Ϯ������ͨ��Ӭ����һ����Ҫ�ص���ֻ��4��Ⱦɫ�壬����������鶯���Ⱦɫ����Ŀ�仯������
������24����˹�Ӭ�ر��ʺ����о������������������֤ʵȾɫ��ѧ˵������ġ�
����Ⱦɫ����ϵ¶��Ŵ�
����19����9O��������Ժ���һ��˼������κ˹��ʱ�ڵ����������ۡ��������µ�����ѧ��Ӱ���£��¸������˹�ͱ���ɭ���ϵ¶����ɵIJ������൱��ij̶����������Եģ�ǿ����ֵ�ͷ��������ǣ���������ͬʱ����һЩ�о��Ŵ������ѧ�ߣ���������Щ����ϸ��ѧ�������ˣ���Ϊ������ϵ¶������������ͣ���ȷ�е�˵����̽���ϵ¶�������������ʻ���������Щѧ�߿�����Ⱦɫ�����Ŵ�����֮����ȻӦ����ij�ֹ�ϵ�������ֹ�ϵ���������е��˶��ܽ��ܡ�Ϊ���˽����ֶ������������һ��ָ���Ŵ�ѧ������ѧ���Ǵӷ�������ѧ�����ġ�κ˹��������ɭ��Ħ����ԭ�������ܶ�����̥ѧ�ġ���Ȼ�ȳ����������֮��������ƺ���һ�ٶ���ǰ���Ժ����۵ľ�����ʤ�������գ���̥ѧ��ȴ��Ȼ��������һ˿һ�Ƶ��ȳ���˼���ر����С�ֻҪ����Ħ�������ڣ�1903�����ϵ¶�ѧ˵�����ۻ�Լ��ѷ�Ի�������۾��ܸо������Ƕ��ϵ¶��Ŀ����Ŵ�ѧ˵�������ǿ��������ȳ����ߵ�ѧ˵�����������顣
���������ǵ��Ŵ�ѧ˵�����������֮�ϵ�ѧ���ǣ����籴��ɭ�Ķ�̬����ѧ˵��theory of dynamic vortices��Coleman��1970������Ϊ�Ŵ���������������������۵�ͳһ���Ϳ���ѧ˵�������������ݡ����ϵ¶��Ŵ�ѧ�Ѿ�ȷ���˺ܾ�֮����һЩ�Ŵ�ѧ�Ҽ�����ࡰ��̬��ѧ˵������R��Goldschmidtֱ��������͵�50�����Ȼ�����Ŵ����ġ��������������Ŵ����й��ɵ�ϵͳ�Ա���Ŀ����ԣ���Ҳ��һ�������Ը��holistiC concept����Լ��ѷ���Խ�������Ϊ����̬�ṹ���ƺ�Ҳ����ͬһ������
�������ǵĶ�����������̬�Կ����Ŵ�ѧ˵�����Ƕ��Ŵ�������������Ⱦɫ������֯������ȴȫȻ���������1890������ڣ������Ŵ������Ⱦɫ��ѧ˵����ʵ�����Ѿ��߱���Ȼ����ʱ��������֮������һ����ȫ��ѧ˵��ԭ���Ƕ��ģ�
������1�����ǿ��ܱ��������ȳ����ߵ�ѧ˵��
������2��û�а����������������Ŵ�����
������3����1885�굽1900���ر�ǿ��ϸ�����ѵĴ����е��˦���档
������4���Դ���ķ��������ر��ע�������Dz��������������Ŵ�ѧ�漰��Ⱥ��������ϸ��ѧ�еĹ��ܷ����������������ġ�
����1900���Ժ��Ŵ�ѧ�ķ�չ�ܵ�һ��żȻ�ɺ���̬��Ӱ�졣�����������̥ѧ������ѷ��E��B��Wilson����ŷ�ļ��ζ������ж�ϸ������ѧ�����˼�����Ȥ���ر����ܵ��������Ѳ�������Ӱ�졣��Ȼ��ʱ����������һЩʮ��ר���Եľ��ж��������ϸ��ѧ�о���ϸ����ϵ����������Ҫ����������ʱ��ϸ�����ر��Ƕ�Ⱦɫ���֪ʶ�����˳�ɫ���ۺϣ�д��ר����ϸ���ͷ������Ŵ�����The Cell in Development andInheritance������1896���ڶ��棬1900������һר���ں���ϸ��ѧ���ϵ¶�ѧ˵���ۺ�������Ļ������ñ�ʲô����Ҫ���������İ�ƪ������ϵ�����£�1905��1912������ƽ��˶�Ⱦɫ����о������⣬��Щ����Ħ�����������������������ã���ΪĦ����ͬ�º�����������Ħ������Ҳ��������ԶӰ�졣�г�ֵ����ɽ�����ѷ��Ϊ�Ŵ�ѧ�����¿�ѧ�Ĵ�ʼ��֮һ��
������Ȼ�в���ѧ����1890����ͱ�ʾ������ΪȾɫ���Ⱦɫ�ʻ���أ�nuclein�������Ŵ����ʣ����ǵ�ƾ��һ�㻹�������Թ�����ʵ�������ݵ��Ŵ�ѧ˵��ֻ�ǵ���1900���Ժ��10���һ��һ�ε�ȷ�����ϵ¶�ѧ˵��ϸ��ѧ2������й�ϵ���Ʋ�ͼٶ��ű�ȷ���֤����������ʵ��֤�������档
����Ҫ������Щ֤�ݻ�֤�����ɵIJ�������ѣ���ΪȾɫ��ѧ˵����ʷ�ͻ���ѧ˵����ʷ������һ��ֻ�����۵ؽ������������жϲ��п��ֱܷ���������ߵ���ʷ�����ǣ�Ӧ��ǿ���������ﲻֻ��Ϊ�˽�ѧ��ԭ�����Ҳ�Ǵ�֪ʶ��չ����ʷ�Ƕȵ����ɲŽ����߷ֿ����ܣ���Ϊ���û��Ⱦɫ��ѧ˵���ȣ������ѣ�������Dz����ܣ���չ��ȫ�Ļ���ѧ˵��
����1900�����·����ϵ¶����ɣ�ʹ��������˼���仯���������������·���������ļ�����������˷dz�����о��ɹ����·��֣�����1880�����1890�����ϸ��ѧ����ͻȻҲ��ʾ���µ����塣�ϵ¶��������Ŵ�����Ⱦɫ��ṹ�����������һ��������Ƕ�������Montgomery��1901��������˹��1992����ɪ�٣�1902��������ѷ��1902������������1902������ͬʱ�����������ɪ�ٺͲ�����Ϊ���ǵĽ����ṩ����ϸ֤�ݡ���Щѧ������ʶ�ؽ�ϸ��ѧ֤�ݺ��Ŵ�ѧ�۵��������Ľ�����γ�������ѧ��һ����ѧ�ƣ�ϸ���Ŵ�ѧ������ѷ����ѧ����������ѧ�ƵĴ�ʼ�ˡ�ֵ��ע�����˹���巶�أ�Sturtevant����������˹��Bridges�������գ�Muller���ڼ���Ħ���о�С��֮ǰ��������ѷ��ѧ����
����Ⱦɫ��ѧ˵
�������Ŵ�ѧ��ʷ��ϸ��ѧ��1900��ǰ��Ľ�չ����Ҳû��ʲô����֤��Ⱦɫ��ĸ����Ժ������Ը���Ҫ��Ⱦɫ����ϸ������֮���Ǽ������ģ���ֹ��ϸ�����˽�������Ϊ��Ⱦɫ�Ŀ�������ϸ˿��ɵ����硣Ⱦɫ������˿���ѽ���ʱ��ȫ�ܽ⣬������һ����˿�������ڿ�ʼʱ�����γɵ��۵��ƺ��õ��������۲��֧�֡���Ҳ����һЩ�о����ϸ��ѧ�������ά����R��Fick��19O5��1907��һֱ���ϵ¶�ʱ����Ȼ������һ�۵��ԭ����ϸ���˾�ֹ��ÿ��Ⱦɫ�屣��������Ժ������Ե��۵�ȷʵֻ�Ǹ������ۣ�����ֱ�ӹ۲쵽��Rabl��1885��������ȷ�����Ⱦɫ��ĸ����Ժ������Եļ�˵������ΪȾɫ���ܽ���ɵ�Ⱦɫ��˿��ϸ���˽��뾲ֹ������һ����˿���ѿ�ʼʱ�����ºϲ���ԭ����Ⱦɫ�塣��ֻ��һ�����ۣ������ݵ����Ϻ��٣����д�����Ǹ���Ⱦɫ�����Ŀ�̶��������������ۡ�Van Beneden������ʮ���¡����Ͳ��������������һ���۵�����Ȩ�������ǡ��������ʲ�����ΪȾɫ�������ѧ˵���������ṩ�˸��߾��������֤�ݡ�����1891�����ͽ����������ǿ���ͨ�����ϸ���˵�ijһָ��Ⱦɫ��ȥ�����ɾ�ֹ�˲�����ÿ��Ⱦɫ�塣������֮�������������Ľ��ۣ������ܾ��ѵ��������ѹ���������ϸ����Ⱦɫ��һ��ض����Ը�������һ������ĸ������1891��410����
��������ϸ���˾�ֹ�ڵ��������Լ�ÿ��Ⱦɫ��ĸ����ԣ��ڽ��쿴��������һ������������������棬Ȼ����1890���ȴ������ˡ�κ��������������Ϊÿ��Ⱦɫ�庬��һ�����ֵ�ȫ���Ŵ����ԣ�Ҳ����˵���Dz������ϵ¶������ϵ�Ⱦɫ��ĸ����ԡ����ǣ����һ��Ⱦɫ��ֻ���и����һ�����Ŵ����ʣ�ÿ��Ⱦɫ��ͻ������Ⱦɫ�岻ͬ��Ҳ����˵���ض����и����ԡ����仰˵�����ÿһȾɫ��������IJ�ͬ���ͱ�����֤�������Ժ����ԡ�
��������Ⱦɫ�������������Montsomery��1901����ɪ�٣�Sutton��1902�����ṩ�˿϶�֤��������ָ������˿���Ѻͼ�����������ЩȾɫ���ǿ��Ը���ֱ�ģ�����ͬһ������Ⱦɫ����ÿ��ϸ�������ж�һ�ٳ��֡����⣬���ǻ�ָ���ڵ�һǰ����������ͬ��Ⱦɫ����ԣ����ᣩ���ڼ�������ʱ�˴��ַֿ��������ģ�������һ���͵ó��������Ľ��ۣ�ÿ�����ֵ�Ⱦɫ���麬�гɶԵ�ͬԴȾɫ�壬����һ�����Դ����ӣ���ϸ��������һ�����������ӣ����ӣ���������van Beneden��1883��۲쵽�����ܾ����γɺ��ӣ���ʼ��������ϸ������ֱ���γ���������ǰ�ļ������ѣ���ЩȾɫ����Ȼ���������DZ�����ͬһ�ԣ���ȫ��ͬ����ɪ�����������½�β�Ľ����ǣ���������ĸ��Ⱦɫ���ϳɶ��Լ�����ڼ�������ʱ�ֿ��������ܹ����ϵ¶��Ŵ����ɵ����ʻ��������ڶ������ֽ���һ˼�����չ����McKusick��1960����
����������Щ�۲첢������ȫ�ų���̬�ϲ����Ƶ�Ⱦɫ��Ҳ���������Ŵ����ʵĿ����ԡ���������Boveri��1902��19O4��ͨ�������IJõ�ʵ��������ֿ����ԡ���һ�־���36��Ⱦɫ��ĺ����в�����ͨ���ʵ�������������ھ��ȣ���ͷ�ĸ���ϸ�����ܹ��õ����и��ֲ�ͬ��ĿȾɫ�����̥��Ȼ����������Щ��̥��ֻ����ϸ����36��Ⱦɫ����ܹ���������������������һ��ʵ�ó��Ľ����ǣ�ÿ��Ⱦɫ����С���ͬ���ʡ���ֻ�е�������Щ����ǡ�����ʱ��������������
���������Ѻ������֤ʵȾɫ�����Ŵ���״�����ͬһ���ɣ�������Ҳ��ʾ�����������������ɪ�ٺͲ����������ػ�����ᵽ����λ��Ⱦɫ���ϣ�ÿ��Ⱦɫ����������Ļ����顣�����ԣ�������ɪ�1903���Ͳ�������1904���������ģ������һ��ȫ���Ⱦɫ���Ŵ�ѧ˵���Ǵ�ϸ��ѧ֤�ݺ��ϵ¶���״����������������۵ó��ġ����ƺ��ܹ������ϵ¶��Ŵ���ȫ������
������ֵ��ǣ���ɪ��-������Ⱦɫ���Ŵ�ѧ˵����������ɪ�ٵ���ʦ����ѷ��1928�������ģ�����Ҫ������ձ������������ȫû�б����ϡ���������ɭ��Goldschmidt�ܲ����ܶ�������һЩ֪��������ѧ�ң���E��S��Russell��Ҳ����1930��ų��ϡ�һ����ԭ����������ѧ˵�Ǹ��ݹ۲��������ƶϡ�Ħ������˵���������ܡ���������ʵ�顱�����Ľ��ۣ�Լ��ɭҲ���������ƵĻ�����ʵ��ɪ��-������ѧ˵���Ǹ���ʵ��ó��ģ�Ħ������ѧ˵�ĵ�����Ȼ���и�����ԭ��
����Ⱦɫ�徭�ɾ�ֹ�ڵ������Ե���1910�����д���֤��֤�������ǵĸ����Ե�֤����Ҫ�Dz�������ʵ�顣�����û����ȷ��֤��֤��ij��������״��һ���ض���Ⱦɫ���йء��Ա�����������������֤�ݵ��Ŵ���״�����֤�������������ͼ��
����Ӥ�����Ա�����ʲô������������������Ǵӹ�ϣ��ʱ���и��ָ����IJ²⡣��������֪�����ڵ��й�ѧ˵���Ǵ���ġ������Lcsky��1950;Stubbe��1965�����е�˵������̥���ӹ�������Ҳ�������е�˵���ɾ���������غ�軹����غ����������е�˵���ɾ��ӵ����������Ի������Һ�ġ��������ȵȾ�������Щѧ˵�Ĺ�ͬ�㣨Ҳ�Ǿ��о��������һ�㣩���Ա������Ŵ��鶨������ȡ�������ܾ������ɺϵĻ������ء���Ȼ������1900���Ժ������Ա���Ŵ�����������ijЩ�������߷�ѧ�Һ��ڷ���ѧ������Ȼ��ֻ��������۴���ʮ��֮�á������ں��滹Ҫ����ijЩ������Ա�ȷʵ�������Ŵ������ġ�
����ijЩ����ϵ¶������߲�û�к�������һ����ʵ����1��1���ԣ��𣩱ȣ�ֵ����һ���Ӻ��ӣ�Aa����һ���������ԣ�aa���ӽ����õ��ı�ֵ��ȫ��ͬ���ϵ¶���1870��9��17�ո��ڸ�������о��Ѿ��ᵽ��һ�㡣����ѧ�ߣ�strasburger�Ϳ���������1900���Ժ�Ҳ�������ͬ�Ŀ�����Ȼ������˹ȴ�������ʵ��֤��ָ�´�������ֲ��к������Bryonia����һ�뻨���Ǿ�������ģ���һ���Ǿ�������ģ������е������Ա�������ִ��ۡ�������������������Ӻϵģ�������ѷ��1910�����������������ӵģ�������ͬ�����ӵġ�����֤��������۳�Ŀ����Ĵ������������ӵģ��������ࣨ�����ˣ���˫��Ŀ���棨������Ӭ��������Ҳ���������ӵġ������������Ա��Ƿ���ܺ�ijһ�ض���Ⱦɫ���йأ����������˴���֤��֤ʵ���������롣
������Ⱦɫ��
������Ⱦɫ���о�һ��ʼ�۲쵽һ��Ⱦɫ��������ϲ���һ����Ȼ��ȫ��ͬ��Henkins��1891��������촻��Pyrrhocoris����������ʱ������һ�뾫�Ӿ���11��Ⱦɫ�壬��һ�뾫�ӳ�����11��Ⱦɫ���������һ��Ⱦɫ��������塣�����ò����Dz���Ⱦɫ�壬Henking��֮����ĸX��ʾ��Henking��û�н���X�����Ա���ϵ������
����������10���з����˸�����������ȷʵ�������Ķ���Ⱦɫ�壬������һ��Ⱦɫ���ڴ�С��Ⱦɫ���ܻ������������������Ⱦɫ���鲻ͬ����Ϊ�Ѿ��۲쵽���ŵ�һ�뾫�Ӿ���X�������ģ�Ⱦɫ���������һ�뾫��û�У�McClung��1901���;ݴ��������µ�������������֪����һ�����ֵij�Ա��Ϊ�����Ψһ���ʾ����Ա�������ƶ��⸽��Ⱦɫ����Ǿ�����̥����ֳϸ������������ʹ���б仯����ϸ����ɸ߶��ػ��ľ��ӵ�Ҫ�أ���Ҳ����˵��Щ���ٲ�Ѱ����Ⱦɫ������Ⱦɫ�壬������Ա�����á�����McClung��һ���۵�ijЩϸ���Ǵ���ġ�NettieStevens��1905����Brush��1978��������ѷ��E.B.Wilson��1905�����þ���ȷ��ȷ������Ⱦɫ����ȷ���Ա��е����á�
�����Ա�����кܶͬ��ģʽ����ʱ�漰������Ⱦɫ�壬��ʱ�������Ӻ��Ա������һЩ������Ӻ��Ա�ȴ�Ǵ��ԡ�������Щϸ�ڿɲο��Ŵ�ѧ��ϸ��ѧ���κ�һ���̿��飨��Wilson��1925��White��1973������Ҫ��һ���ǣ���֤��һ�ֱ�������״�����Ա𡢺�ijһ�ض���Ⱦɫ����ϵ��һ��
�������Ƕ�������ϵ���״ξ�����֤���������������д�����Ŵ�ѧ�о�����Ϊ����֤������״����Ⱦɫ�������Ⱦɫ�壨��Ⱦɫ�����ͨȾɫ�壩֮��Ĺ�ϵ�������о���ʵ��Ⱦɫ���Ŵ�ѧ˵�����ͷ�˾���Ħ�������ǵ��о�������һ��Ⱦɫ�����Ŵ������϶�����ͬ��ѧ˵����һѧ˵��190o���Ժ�һֱ�����У���Ȼ��ʱ�Ѿ����ֲ������ֵ�Ⱦɫ��Ĵ�С����һ�¡�19���Ͱˡ���ʮ�����һЩ����ѧ��֮���Լ����һ����������˵�������ŵ�ѧ˵�ܿ�������Ϊ����Щ���������е�Ⱦɫ���ȷ�����ơ�
������Ⱦɫ��ĸ����ԣ������ԣ��Ѿ��õ�֤ʵ���������ٷ�����һ����״���Ա𣩺��ض���Ⱦɫ���й�֮���Ŵ�ѧ�ұ���Լ�������̽��������Ĺ���Ⱦɫ�����״�����⣬������Լ��ѷ�Ļ��������˵����Ⱦɫ�������֮��Ĺ�ϵ���⡣Ⱦɫ���Dz�����Ϊһ������������ȫ����״���ȷ�˵����һ���¿���ս����ָ�����ģ�������Ļ����Dz���λ��Ⱦɫ����ض���λ�������ͬ�Ļ�����ͬһȾɫ���λ�ڲ�ͬ��Ⱦɫ���ϣ�����֮������ϵ��������Щ������һ���Ƚ϶̵�ʱ���ڣ�����������1905-1915��10��֮�䣬ʵ���ϴ�����1910�굽1915�꣩��ȡ���˴𰸣���Щ����ͨ��������Ŵ�ѧʵ�鲢������ϸ��ѧ֤���������ȡ�õġ���Щʵ�����Ǵ��൱���ϵ¶�����ʼ��
����Ħ����1909�꿪ʼ������Ӭ�����Ե¸�����¼���ͻ���ӡ������ͼͨ����ͬ�Ļ�ѧҩƷ����ͬ�¶ȵĴ����Լ��ƺ�X�߷��������������Ĺ�Ӭ������ͻ�䣬���Ƕ�δ�ɹ���Ȼ�������Ĺ�Ӭϵ��������Pedigreed cultures������һ��������Ӭ�����ں��۹�Ӭ��������Ⱥ�С�
������һ������ʵ����ʵ������������г����˵����쳣�ĸ��壬ȴ������һ��������ʵ��ѩ��ʽ���о��ȳ�����������������ǣ���������ۡ���״�����������ġ�����һ�����İ����۹�Ӭ��ͬһ���Ĵƹ�Ӭ���䣬Ħ��������ȻF1�����Ǻ��ۣ�����F2���ֳ��ְ�����Ӭ�����ʾ���۵��Ŵ����������ԣ�����Ȼ���ɺ��ۻ�����Ȼ�仯�������ġ�Ħ���ڼ���ǰ�����ʹ��¸����ں�����ʵ���ң�Ҳ��Ϊ���µ�λ�������Դ�����˵¸���ġ�ͻ�䡱��һ������ڵ¸���Ľ���ͻ��ѧ˵�Լ��¼���ͻ���Ⱦɫ�����ʣ���һ�����ת�������˲����ĺ���������Ķ���ʮ���������һ����˼����ң�Anen��1967��Mayr and Provine��1980����Ȼ���Ŵ�ѧ�Һͽ����������ϰ����Ħ�����衰ͻ�䡱��ʵ��º��塣
����������ѧ����ʷ�ϻ���������Ħ��������ͬ��һ���������������䡣��˺���ȷ��Ħ��ʵ���ҵĴ����о��ɹ����־���Ӧ�鹦��˭����Щ��ʷѧ�������ڽ��������еĹ�����֮������ѧ����ͬ�¡���δ��̫���ݡ�Ӧ����ס�Դ�Ħ����191O��7�·������Ĺ��ڹ�Ӭ�о��ĵ�һƪ�����Ժ��������������������13ƪ�й�20�����Ӭ�����������ԣ�ͻ���͵ķ�������Ϊ�����¡��ڷ��ְ��ۺ��ַ�������������������ͻ���ͣ����ᷢ����ȫ��������ɫ��ɫ������������Ħ���ں���Ͳ������ϵ¶��Ŵ��Ĵֻ��ƣ���Щ���������������Ĺ��ס��������գ�Muller��1946����˵�ģ������۽�����������д���������۹�Ӭ�о���ʼ�ε���ʷ��Ħ���Խ����������֤�Լ����������Զ����Ҳ��Ƶ���Ĺ۵����һ�����ף�����Ҫ������������ϵ¶�ѧ˵�ķ��֣���һ���Ȼ�Ծɻᱻ���ϡ�������������Ҫǿ������Ħ�����˵����ض������ͽ������������Ĺ��ף���Ϊ�Ժ�������ص���������϶����ǹ�Ӭʵ������ÿ���˵�����ס�Ħ��������ͬ���ڸ��ױ��Ǵ�ѧ�ġ���Ӭ�ҡ��������˳�ǧ����Ĺ�Ӭ����������ϸ�����Щ��Ӭ�з������µ�ͻ�䲻�ϳ��֡�Ħ���ܿ죨191O-1911�궬���Ӹ��ױ��Ǵ�ѧ��δ��ҵѧ������ѡ��˹���巶�أ�Alfred H,Sturtevant���Ͳ�����˹��Calvin B��Bridges�����˵�����ʵ���ҹ������������գ�H��J��Muller��Ҳ�μ�������о�С�飨������Ħ��ָ���¹���ѧλ�������С�������Э��������ѧ�е�һ����ʷ�ѻ������ڿ�ѧʵ�����й�ȥ��������˼������ĵ����պ���˵ij������顣����Ҫ������Ħ�����˵�̬�ȣ�������������о����ں���һ�𣬴�ȡ��ؽ�����������Ĭ�С���Stuvtevant��1959;1965a ����
�����ڶ̶̵ļ���֮�ڴ����Ŵ�ѧ��һ����Ҫ���涼��Ħ��������С�������������DZ���ɭ���¸������˹����������Castle���Լ��������ڵ��ϵ¶���������û���ҵ�����ȷ�𰸣�ʵ������û�������ȷ�����⣩����Ħ��С���ɫ������ˡ���������ˣ�һ����Ҫ��ԭ����Ħ����Ȼ����̥ѧ�ҳ���������������������ѧ���巢����������һ�ߣ���������עȫ���ڴ����Ŵ�ѧ����û��ȥ�ܲ��Ŵ����ɶ���̽����ʵ����Щ��ʵ����Ŀ��ܽ��͡�����һλ���ľ��������ߡ�
���������
�����ϵ¶�����˽��������״�dz�Ⱥ���֣�������ѡ�õ���״�����dzɶԳ��֡�1900���Ժ���о�����֤ʵ�˶�Ӧ��ij��������״�����ʻ��������ж�����һ�ı�����ʽ������ֱ֮���ϣ���ģ���Щ����һ�ģ������������Ӿ��ǡ���Ի���allelomorphs������ɭ��������λ����alleies�����ϵ¶��Ŵ���������һЩ��������״�Ķ���һ���ӵķ��ֱ�Ա��������������������ȫ�µĽ��͡���ʹ���������㶹�Ĺ⻬����Ƥ�������̣��������������Ƴɶ���״��������ͬ�����ʻ�������λ��������һ�ֵ���״Ӧ�������ͬһ�Ŵ����ʵ������뱾��
����1904�귨������£�Cuenot���ڼ����з���һ����״�������������ϵĵ�λ���������ڼ�����һ�����������Ƥɫ�����ǻҡ��ơ���ɫ������ɭ����������Shull��Ħ���Լ������Ŵ�ѧ�Һ���Ҳ���������ิ��λ�������������ABOѪ�;���������֪�����ӡ�Sturtevant��1913�����Ƚ����˸���λ��������֮������ͬһ������λ���ĸ��ֿɹ�ѡ��״̬������ȫ���˱���ɭ�Ļ������õ��л���ѧ˵��Presence��absence tbeory������ijЩ���������һ��������50�����ϵĵ�λ��������ţ��Ѫ�ͻ����Լ�ֲ���ijЩ���Ի���compatibility genes�������������֯���Ի����ϵ¶������������ijһ��������Զֻ�е�����λ�������ܾ�ʱ���ܹ��ʹ�������Ⱥ������е����ͬ���κ�һ����λ�����ϡ����Ŵ�ѧ��ʷ�Ϻ�����������ijЩ�ӽ��л������Ϊ���λ�������ƶ��������ӽ�������ˣ����λ����pseudoallelism����Lewis��Green����������ķ���ʹ���ǶԻ����ʵ���ʶ��������һ���������ģ���
����Ħ���о�С��Թ�Ӭ�İ��ۻ����Լ���Ӭ������ͻ���͵��о���ȷ��֤����һ���������ͻ�����һ����λ����������ͻ��ɵ��������ĸ���λ����ͬ��ֵ��ע�������Щͻ�䲽���ǿ���ģ����۹�Ӭż��Ҳ�ܲ������ۺ����Ҳ������Ҫ�ķ�����ij������һ���������µ�λ�������µĵ�λ�����һֱ���ֲ��䣬��������֮һ��������ͻ�䡣��˻�����ص��ǣ����ļ�����ȫ�ȶ��ԡ�����ͻ��ķ��ֲ������˻ص���ʽ�Ŵ���ǡǡ�෴����֤ʵ���Ŵ������ǻ����̶�����ġ���������˵������Ӳʽ�Ŵ��ľ������������������Ŵ�����ֱ�����ڵ��ȶ��ԣ�Ȼ�����־��н����ݱ���������ͻ��������
�����ܿ��֤ʵ��һ���������ﶼ��ͻ�䣬��������������鶯�ﵽ��Ķ������ֲ����������ʵ�ϴ�1920��ֱ��195O���о�ͻ���ƺ�������ϣ���ġ��ܲ����Ŵ����ʱ��ʵ��о�·�ߡ�ͻ����̵��о�ͬʱҲ�����һЩ���ڽ������⡣��ͻ��ʱ��������ʲô�仯���ܲ����ڿ��������£�����ʵ�������£��������죿�¸�������1904����ᵽ��X����������߶��ܴ�����ϸ�������������ı���ֳϸ���е��Ŵ���������Blakeslee��1936�����Դ�1901���Ժ�һֱ��ͼͨ��X���ߣ������ߣ��¶���䣨�¶��ݿˣ�����ѧҩƷ�Ȱ취�շ�ͻ�䡣���ڸ��ּ����ϵ�ȱ�ݣ���Դ�����ʣ���������С�ȵȣ��������û�еõ���ȷ�����ͨ�����հ��۲��ӺͶ���������עȫ����Ѫ�����о�һֱ��1927���ȡ�óɹ���
������״�������������
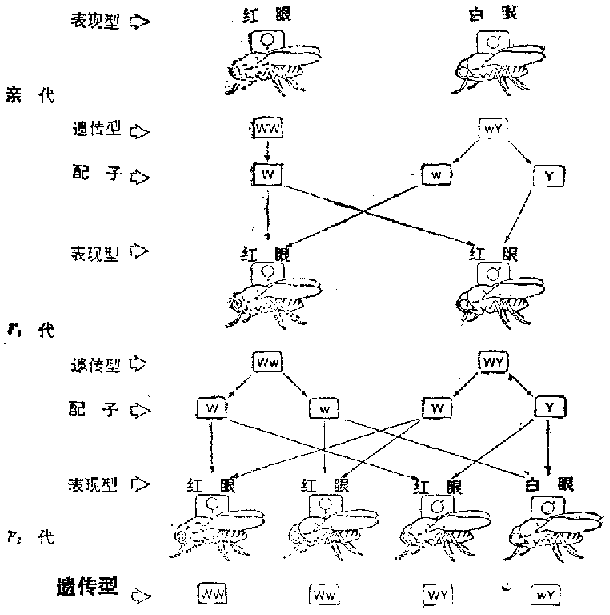
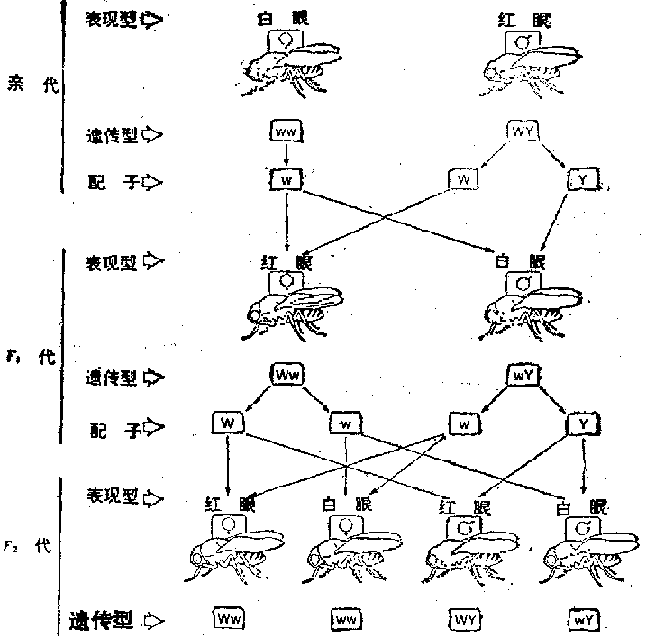
�����ϵ¶�����Ҫ����֮һ�ǡ����֣���ϣ��е�ÿһ�Բ���ͬ��״����Ϊ�������ױ�ֲ����������������ء���1866��27������������һ�㽫֮��Ϊ��״������϶��ɡ����磬�ϵ¶�������Բ�λ�ɫ��������������״�������ԣ����㶹Ʒϵ��������̶��������߶������ԣ�����һ���㶹����Ʒϵ�ӽ������ڼ����ɵõ�Բ�ƶ��������̶����ı�ֵ����3��1��������ر����������ܹ��ɵ�556������������315��ΪԲ�ƣ�101����ƣ�108��Բ�̣�32�����̣���ֵ�ӽ�9��3��3��1�����ÿһ����״��Բ���壬�ƶ��̣��ı�ֵΪ3��1��Բ����Ϊ���ԣ���������������״ȴ�DZ˴˶������з��롣�ϵ¶���������������������״Ҳ����ˣ����һ����Ϊ������״�������һ������϶��ɡ�
�������ϸ����ֻ��һ��װ���ɶ�ѿ�Ŀڴ����������γ�֮ǰ��Щ�ɶ�ѿ����ֿ����������䣬�ϵ¶�����һ���־ͺ���Ȼ����ֵ����֡�Ȼ������ϸ������������֯��Ⱦɫ��ģ���Ⱦɫ���������γ�ʱ������Ϊһ��������з��룬���ԾͲ����б�Ⱦɫ����Ŀ����Ķ�����״�ԡ��ϵ¶����߸���״���������һ�����ʵ�������Ϊ�������˺ܾ�֮�����㶹ǡ��ֻ���߶�Ⱦɫ�壨�����ģ�
�����Դ����·����ϵ¶����ɺ������Խ��Խ����ӽ����飬����Щ�����з�������������ϲ���������⣨�����ǿ���˹��1900�����������з��ֵģ����������ɱ���ɭС�鷢�֣���Ȼ���������漴���ᵽ��ԭ������������ѽ��͡��ڰ��۹�Ӭ���Ա���۾���ɫ�������������һ����ܿ�ͱ�Ħ�����֡�������F1����Ӭ�ӽ�ʱ��F2����Ӭ�к�������۵ı�ֵΪ3��1�������еİ��۶����۹�Ӭ���������еĺ��۹�Ӭ�д��۱�2��1����ͼ2a������������һЩ�ӽ��У�������������ϡ�������۴ƹ�Ӭ�������������۹�Ӭ�ӽ�ʱ�����еĴ��Ժ�����Ǻ��ۣ����е����Ժ�����ǰ��ۣ�ͼ2b����
 |
����ͼ2a����ĸ����F1����Ӭ����ĸ��XȾɫ��Ϊ���ۡ����������ϵ¶����룬50����
F1��Ӭ��F2��Ӭ��Ϊ����
 |
���������ԣ��Ի�����۵���ɫ����������ϡ�
����Ħ����1910����������۲�϶��۾���ɫ���ӣ��Ӻ�ɫͻ��ɰ�ɫ���Ǻ;����Ա��X���ӻ���ż���ġ�һ���Ժ�1911��384�����صش�Ⱦɫ��ĽǶ�������������״ż���������Ƿ��ֵIJ����ϵ¶������ϵ�������������Ⱦɫ�����ദ�ܽ��ġ��������ϡ���association of factors����ϸ��ѧ�ṩ��ʵ��֤������Ҫ�Ļ��ơ���������һЩͻ�䣬�������ɫ��С�γ�Ҳ�ǣ����Ա������ģ�Ҳ����˵λ����Ⱦɫ���ϡ���������״����Ⱥ���Ա��أ���Ȼ�Ǵ��ڹ�Ӭ������Ⱦɫ���ϣ���ЩȾɫ�屻��Ϊ��Ⱦɫ�����ͨȾɫ�壨����Ⱦɫ�����ֿ�����
�����¸������˹����������ɪ��ʵ����Ҳ���Ѵ�������Ԥ�Թ��������������Ǹ���Ⱦɫ��ĸ����Ժͣ�ͨ��ϸ����˿�������ڵģ������Խ��������ġ�
����������
����������˹��Ħ��С��ij�Ա֮һ����1914��ΪȾɫ���Ŵ�ѧ˵�ṩ��һ������˵������֤������һ�����۴ƹ�Ӭ��������XȾɫ���϶���������ɫ��λ����һ�����������۹�Ӭ�����䵥��XȾɫ���ϴ������Ժ�ɫ��λ�������ӽ�ʱ��F1��������ͬ��Ŀ���Ӻ��Ժ��۴�Ӭ�Ͱ�����Ӭ�����������ױ����Ŵ�����������ģ���Ȼ��ˡ�������Ħ����ʵ�����г�����һ���쳣�Ĺ�ӬƷϵ�����������ӽ���������һƷϵ�Ĺ�Ӭ����F1����Լ��4��3���ǰ��۴�Ӭ�ͺ�����Ӭ�������������ϸ�ںͽ��ͣ��ھ����Ŵ�ѧ�̿����ж�����ϸ���ܣ����ﲻ���ظ���������˹��������һƷϵ�Ĵƹ�Ӭ����������XȾɫ�廹��һ������YȾɫ�塣��һXXY�ƹ�Ӭ�����ǵ�һ����������XXȾɫ����쳣���ӣ����ڼ��������еIJ������һ��Y�����ھ�������ġ���������һ������������Ⱦɫ�壨����X��һ��Y�ĸ������γ�����ʱ������������XȾɫ�������ͬ�����ӣ����ӣ��У��γ�X��XY���ӣ�ʵ�������γɵ�������91��8��������������������������X����һ�����ӣ�Y������һ�����ӣ��γ�XX��Y���ӣ�Լռ8��2���������������ġ�����X��Y�ľ����ھ���XXX��YY���Ӳ��������������Ȼ�������������������Ӭ��XY���Ͱ��۴�Ӭ��XXy����������ͼ3����������˹�����������ϸ��ѧ�۲�֤ʵ����֤ʵ����һƷϵ�Ĺ�Ӭ��XXY��Ӭ��XYY��Ӭ���ڡ��ڴ���ǰ��Wilson��1909�����Ժ��Ⱥ�����һЩ���������������庬�ж���ij�Ⱦɫ�塣����������������21��Ⱦɫ�壬����Ϊ���ڲ������21��Ⱦɫ�����壬��������������֢��Down's Syndrome����׳գ���ԭ���ںܶ���ֲ�����еĸ�����ж����Ⱦɫ�壨���壩��ʧȥ��һ����Ⱦɫ�壨���壩����Щ�����ܹ����沢�������о�ͬһ����ĸ���ƽ��ЧӦ��������������12��Ⱦɫ����κ�һ�����岻���ܹ��������������̬���̲���23��Ⱦɫ�����һ�ֵ���Ҳ����ˡ�
����
 |
����1902���Ժ���Ȼ��һЩ����ѧ������̸��Ⱦɫ���Ŵ�ѧ˵����������ȷ���徿����ʲô��ʱ����ʮ��������������ָ�����㣨Roux��������������������ͬ���Ŵ����ӣ�����Ӧ��˵������Ⱦɫ���ϵ�ֱ�����С�Ȼ���Ⲣ���������ȫ����ϸ��ѧ����1870�����1890�������������϶����Ŵ��йص�Ⱦɫ������1900���Ժ����Щ���������ϵͳ�о���������Ħ���о�С����1910���Ժ���о�������ٽ���Ⱦɫ��ѧ˵�ķ�չ��
���������Ⱦɫ���������γ�ʱ����Ϊ��ʼ���ܡ����ӵ�ϸ���˺;��Ӷ��ǡ������塱��Ҳ����˵���Ǿ��е�Ⱦɫ����Ŀ�ǣ������壩��ϸ��Ⱦɫ����Ŀ��һ�롣�������γ�ʱȾɫ����Ŀ����������һ��Ķ�������������Ӱ���Ŵ���
�����������ѣ�Reduction division��
����Van Beneden��1883�����ֵ��׳�������ܾ�ʱ�۳澫�ӵ�����Ⱦɫ�������ϸ���˵�����Ⱦɫ�����ϣ��γɾ����ĸ�Ⱦɫ��ĺ��ӵ���ϸ���ˡ�����ܾ������ϸ����������������������ÿ��ϸ���������ĸ�Ⱦɫ�壻��ÿ��ϸ���ǡ������塱����Ⱦɫ����Ŀ�����ӵĶ����������ÿ���ܾ���Ⱦɫ����Ŀ������һ�������Ӵ���Ⱦɫ���������״���һ�������һ����ÿ��ϸ���ܿ�ͻ���г�ǧ�����Ⱦɫ�塣�����ԣ����ܾ������б�Ȼ��ij�ֹ��̵���Ⱦɫ�������ijɱ�������Strasburger��1884����κ˹�����Ⱥ�����������γ���ǰ������С��������ѡ�����������1887��1888��Ҳͬ��κ˹�����������������ά����1890�����������ѹ��������꾡������ȷ��������
����ϸ��ѧ�߷��ֶ������γ�����ʱ��������������ϸ�����ѣ����Ǻ���������˿����ʮ�ֲ�ͬ��������֮��Ϊ�������ѻ������ѣ�me1os1s�����ڼ�������ʱ����������ʲô�ջ������߸�ȷ�е�˵Ⱦɫ�������������ٵģ��������ȴ�������۲������������ʱ�ܿ�ͱ���ҹ��ϵ�Ψһ��ʵ����ĸϸ���;�ĸϸ����������������Ӻ;��ӵ�ϸ������Ⱦɫ����Ŀ��һ�����ϸ����ͬ�����Ƕ����壻���ɼ������Ѳ��������ӣ����Ӻ;��ӣ���Ⱦɫ������ֻ��ԭ����һ�룬�ǵ����塣���������ѵ�ʵ�ʱ�����˽����Ժ��֪����ϸ����ϸ���˵ij���;��ӵ���ȫ���ƣ�����էһ��������ϸ���ij������ȴʮ�ֲ�ͬ����������潫֮�ֱ���ܡ�
����ÿ��ϸ���˵�Ⱦɫ��������һ��ͬԴȾɫ����ɣ�����һ�����Ը�������һ������ĸ�����ڵ�һ�μ�������ʱ����һ��ͬԴȾɫ��˴˴Ӳ������úܽ���������ϳɶԵĹ��̳�Ϊ�����ᡱ�������������Թ�����������һ��ϸ����ȫ����������߱˴����ϵ�������У����������۲������ȷ���ڵ�һ�μ������ѣ���ǰ�ھ���������һ���¡������ֹ���30���ͨ���Ŵ�ѧ����������̻����о�����������ģ����������ͺ���ά�����۲쵽�����ձ��ڣ�Ũ���ڣ�֮���Ⱦɫ��ֿ�������������˿����ʱ����������ϸ��������������ѵ����ܽ��У���һ����Ⱦɫ�壨daughter chromosomes���Ե����������ʽ���ų���
�����������ѵĵڶ���ϸ���˷��Ѻ����⣬��Ϊֻ�Ǻ˷��Ѷ�Ⱦɫ�岢�����ѡ��������һ��Ⱦɫ������Ӻˣ���һ�������һ��ϸ���ˡ��ڻ׳棬Ⱦɫ����Ŀ��������ĸ����ٵ���������ڶ��η��ѱ�κ˹����Ϊ�������ѣ�reduction division��������ϸ���е������Ӻ�֮һ����Ϊ�ڶ������屻�ų���
�����������γ�ʱҲ����ͬ�������κ˷��ѣ�����ͬ����û���Ӻ��Լ�����ʽ���ų����������γ���һ���ĸ����ӣ������Ǵӽ����˼������ѹ��̵ľ�ĸϸ���е�һ��ϸ���˲����ģ���һ�����ά�ˣ�1890������ָ�����������ڵڶ��μ���������Ⱦɫ����Ŀ��û������һ�����������ĸ����ӵ�Ⱦɫ����Ŀͬ��Ҳֻ�Ƕ��������ϸ����һ�롣�������������ܵ�ֻ�Ǽ�����������������̬�������ͣ�ȫ�����̵�ϸ������VanBeneden������ά�ˣ�κ˹���Լ������˵ķ��ֺͽ��������ɣ�Churchill��1970�����Ͷ���һ�������ʶ��չ�����������ɵIJ��������������ά�˺�κ˹���ڽ����ϱ˴��������ܴ�����������Ҫԭ����ά�˶��������ں��Ŵ���������ĸ����Ⱦɫ�����ܾ�ʱ�˴��ںϣ�����Ϊ��ÿ��ϸ�����Ѻ�Ⱦɫ�屻�ܽ⣬����һ��ϸ������ǰ��������Ⱦɫ������ɡ��͵ؿ�����������ų�������Ⱦɫ���ʵ����ļ��١��෴��κ˹������Ϊ�ܾ�����ĸ����Ⱦɫ��һֱ�ֿ���������Ϊÿ��Ⱦɫ������˿���ѡ����������Լ����ľ�ֹ�ڵ�ȫ�������ж����������ԡ���������������κ˹�������������֤������ȷ�ġ������������еó�������Ҫ�����ǣ������ų��ļ���������Ⱦɫ��ĺ����ʹ�������ϸ�����е���ȫ��ͬ��������γɴơ�������ʱ��ϸ���˷��ѣ��������ѣ�Ҳ��ȫ��ͬ�������ڴ�������������γ�һ���Ѻ��������壬�����������γ��ĸ����ӡ���Ȼ����ά�˺�κ˹���������۵���ϸ��ѧ����ļ�����ϸ�ڣ�������Churchill��1970�����м��ص�ָ����������ʵ����ȴ�������ǵ����������ͬ�������ġ�����ά�˴�������������������ѧ��Ӫ��κ˹���������̬ѧ�����۷���ѧ�ɡ�
��������Ҳ�������Ŵ�ѧ������������������ѵ�ϸ��ѧ���ݡ��ٶ��º��ӵ��γ������ڸ�����ĸ��Ⱦɫ�����ϵĽ��������һ��Ⱦɫ����е�λ����A�����Ը�������һ�����е�λ����a��Ⱦɫ�壨����ĸ������ƥ���������Aa��������º��ӵĵ�һ�����ѿ�ʼ��������ͬԴȾɫ��ͽ�һֱ������ԣ��ڷ����е������������ϸ���������Ӻϵ�Aa��ֻ���������γ�ʱ�ĵڶ��γ�����ѣ��������ѣ�ʱ������ͬԴȾɫ��ŷֿ����γ�ͬ����Ŀ�ľ��л���A�����Ӻ;��л���a�����ӡ�������һ��Ⱦɫ����뵽��ϸ����ȥ����żȻ�������ģ�������һ���ϵ¶��ķ��붨�ɾ��ܱ��ܾ����ú������γ�ʱ���۲쵽��Ⱦɫ����ΪԲ���ؼ���˵�����������ѽ��ܵ�������Ժܺõؽ��������ͷ��������ǵ��ˣ����û�н����������ܽ���ǰ����ָ���IJ���ȫ��������������ü��������е���һ�ֹ��̡�������˵����ֲ��ļ������Ѿ�Ⱦɫ����ԺͶ������ͬ����ͨ������������ʷ��һ����ͬ�Σ������γ�֮ǰ����
��������
���������κ��������״��������Щ��״���Ŵ�����ԶԶ����������Ⱦɫ�����Ŀ�����Դ�һ��ʼ�����ˣ�Correns��1902��Sutton��1903������Ϊÿ��Ⱦɫ��ض�Я���м���������������ࣩ������һ��ܿ�ͱ�Ħ���о�С��֤ʵ��Ȼ������Ⱥ��ÿ������Ⱥ��ij���ض���Ⱦɫ���й������ķ���������������⡣���һ��Ⱦɫ���ϵ����л����ܵ�������һ����ôһ�����ォ��ֻ���������е�Ⱦɫ�����Ŀ������Ķ����Ŵ���λ����ͻ������ʩ�Ӽ����Լ�����¸��1903�����о�F2������ʱ���ó��Ľ����ǣ�F2���ֵ�����Ŀ��ܻ���Զ������ȫ�������Ĺ۵㡣���������ڼ������ѵĵ�һǰ���гɶԵ��ױ�Ⱦɫ�巢������λ��������exchange of units�������ڡ��ж��ٺ���һЩ����λ���ύ��������ܴ���żȻ����1910��243������������������������������Ҳ��Ԥ�Թ����ཻ����1904��118�����Ŵ�ѧ�����ܿ��֤����ͬһ��Ⱦɫ���ϵĻ��������Dz���ȫ�ġ�����ɭ��Saunders��Punnett��1905�����Ƚ���������۲��о��������㶹��Lathyrus�����������֣���ɫ�ͻ�������״�˴˲�ͬ���ӽ����F2�У����Ǽ�û�еõ�Ԥ�ڵ�9��3��3:1�ı�ֵ��Ҳû�з��ּ�3��1��ֵ��ȴ������69��5����˫���Ը��壬19��3��˫���Ը��壬�Լ�����5��6���Ӻ��ӡ���Ȼ��������״�Ļ���Ȳ���������ַ���ȫ��������11�����⣩������ɭ�������һ���ر��˵��������һ����������������Ⱦɫ��ѧ˵������Ҳ��û�п��ǽ�����
������ȥ�������������ϵ¶�û������������������һ�����¡��㶹��Pisum sativum��ֻ��7��Ⱦɫ�壬�ϵ¶�Ҳ�����о���7����״������û�����������Dz��������ôӶ�Ϊ�ϵ¶�������һ�������鷳����������������������֪���ϵ¶�����ʽ���������㶹����֮ǰ�������˼���ʱ�����Ԥ���ӽ����飬���ܿ��ܽ��ڼ�������ʾ������ϵ�һЩ��״���������dzɶ���״�е�һ����״�����ò��ˡ�Ҳ����������֮ȡ��������ϵ�������ƫ��������ϵ���Щ��״�����ijЩ����Ļ���ͼ�ࣨ�룩�㹻��ʹ�Ƕ�����ͬһ��Ⱦɫ����Ҳ����ʾ�����������
������Ħ���о�С�鿪ʼ��������������Ӭ���Ŵ��ṹʱ����������ȫ���������������Ϊһ���������⡣Ħ������ͬ�·��֣����������ķ��Ⱥܴ���ʱ���Ե���1%�������ı������������ͣ�
������һ���������Ϊ������һ���������Ի���-�ƣ��壩ɫ��y�������ۣ�w����С�ᣨm��-λ�ڹ�Ӭ��XȾɫ���ϡ����һ�������������������Ӭ��һ�������ƹ�Ӭ�ӽ�������Ԥ�����������Ի���������Ⱥ����ʽ��F2���г��֡�ʵ���ϻ�ɫ�Ͱ��۵������ڹ�Ӭ����1��3�����ѣ�����С��������32��6�����ѣ���ɫС��������33��8�����ѡ�����������Щ���֣�
������Щ�����������ֵͨ�����ǰ��¸�������ĵ�λ����������������żȻ���������͡����Ǹ���1900������ڵ�ϸ��ѧ�о�����������ͬ�Ĵ𰸡��ԴӲ������ͺ���ά�˵Ŀ����Թ����Ժ��20��Լ�������ϸ�ڵ��о���ȡ�ó����չ���ڵ�һǰ����Ⱦɫ�壨Ⱦɫ���ʣ��ı仯���ٿ��Է�Ϊ6���Ρ�������һ���������ɶԵ�Ⱦɫ����Ȼ��ϸ������ÿ��Ⱦɫ���ѷֿ���Ⱦɫ��˿��Ⱦɫ���壩������ν�����ڡ�������Ⱦɫ����˴˷��������γɲ�״����
��������ʱϸ��ѧ��Janssens��1909��ָ�������ĸ�Ⱦɫ����˴�������һ��ʱ��һ�������ĺ�һ��ĸ����Ⱦɫ������������DZ˴˽��洦���ѣ���ͷ����������ʱ���Ǹ�������Ķ�ͷ����ĸ������Ķ�ͷ����������Ҳ����ˡ�������Ⱦɫ������Ȼ��������������һ�����γ���һ�������桱��chiasma�����ڼ������ѵ�һǰ�ڵĺ��ڽ��dzɶԵ�����Ⱦɫ�屣�ֽӴ��ĵص㡣��Janssens�Ĺ۵㽻���ʾһ�������ĺ�һ��ĸ����Ⱦɫ��Ľ��������������һ���ɸ�����ĸ��Ⱦɫ���Ƭ����ɵ���Ⱦɫ�塣Ħ��С�����о��IJ���ȫ������Janssens�Ĺ۵������
����������������˸������²���30��ʱ�������ܾ�����һ�ֽ�������ȷ�ģ���Whitehouse��1965������ֵ��һ����һ���飩��Ȼ�������ѳ��֤ʵ���������������ڣ��漰�ĸ�Ⱦɫ�����е��������塣���֤ʵ��������һ��ʼ�ͷ���������Gred��1978����
����Ħ������������˹���巶��LewiS��1961����ΪԴ�ڽ����IJ���ȫ�����ķݶ��ʾ�Ŵ�������Ⱦɫ��������λ��֮��ģ�ֱ�߾��롣Ⱦɫ������������֮����ѵĻ��ᣨҲ�ǽ����Ļ��ᣩȡ����������������Ⱦɫ���ϵľ��룻�������������ѻ������١�������һ����˹���巶�أ���ʱ���19�꣡�������˻�����Ⱦɫ���ϵ�λ�ú�˳���Ƴ�����ͨ��Ӭ��Drosophilamelanogaster����XȾɫ��ĵ�һ��Ⱦɫ��ͼ��������1913�꣩������֤֮ʵ�˵�ʱ��֪������һȾɫ���ϵĻ���������Ⱦɫ�����������еġ�
���������ڵ�ʵ������һЩ����ì�ܡ����գ�Muller��1916��ָ����һ����Ⱦɫ���Ͽ��ܷ���˫�������ҽ���Ĵ��ڽ�������Ⱦɫ�����뽻���ڽ����Ľ�һ�����������ǵ��������·��ֵ�����˫��������ţ����ų�������ì�ܣ�Ħ����һЩ����������������Щì�������ɽ���ѧ˵����ȷ�ԡ�
�����Ŵ������Ⱦɫ��ѧ˵�����Ѿ������û���ѧ˵��Morgan��1926�������䡣1915��ǰ��Ħ������ͬ���о���һ�ٶ���ͻ���ͻ������Ƿֳ��ĸ�����Ⱥ����Ӭ���ĸ�Ⱦɫ��dz�һ�¡�����Ⱥ��Ⱦɫ��ʵ�ʵļ��֤�����˱��������Ȼ��һֱ��1931��Stern������ijЩ�쳣����X����Ƭ�������ڵ��ĸ�СȾɫ���ϣ�Ϊ�����ṩ��ϸ��ѧ֤����ͬһ��Creighton��������пˣ�1931����ֲ����ף�����Ҳ���������֤�ݡ��������׳���ϸ���Ŵ�ѧ�������о����ϡ���Ȼ��û�к����ڹ�Ӭ�о��зdz������ľ���Ⱦɫ�壬�����������е�ȫ��ʮ��Ⱦɫ������̬�϶���ͬ��������ʱ���ж����Ⱦɫ����ڡ�������п�����������Щ�ص������30��������ֳ�ɫ���о������ͻ�������ã���һ������Ȼ���ݷḻ���־��ж�������ȴһֱ���˺ܶ����ڷ����Ŵ�ѧ�ҵõ�����ͬ����֮��ű��������ϡ�
�������������ܵ��йؽ����������ʷ�ǹ��ڼģ�©���˺ܶิ�����⡣���罻�棨�ɽ���������Ⱦɫ���Ƭ֮�����������ʵ��һֱ�������۲��ݡ�ÿһȾɫ����ϵĽ�����Ŀ����һ�£���ʵ����ijЩ����²�û�н������������۹�Ӭ�����ڵ�һ�μ���������Ⱦɫ���ȷ�и���ʱ���Լ��γɽ��棨Ⱦɫ������Ѽ��K�ϣ���ȷ��ʱ�䣬��������Ĵ����Ƿ����DZ�ʾ���������кܶ����顣����Ҫ���ǣ�Ⱦɫ���в�ͬ��Ⱦɫ�������Ϊ������˵��̳Ī��һ�ǡ������Ϊ���ͽ��������Janssens��Ħ���Ķ��Ѳ���ѧ˵��breakase��fusion theory��������ijЩѧ�߽��ܣ�����Belling����������ˡ�����ѡ��ѧ˵��copy-choice theory����Winkler���������ת�䡱ѧ˵��gene��con��version theory������Ȼ������ѧ˵���û�еõ����ϣ�ȴ��ʹ���ǽ����˴�������Ӷ��Խ��������Լ�����ı������˸�������˽⡣���ڻ�û��������ѧ˵�ıȽ��о���ʷ����������Щ�����ϵ���ϸ����������ϸ��ѧ���Ŵ�ѧ�̿��顣������Grell��1974������Ҫ���ǣ�һ�п����������������ն��ܰ�����Ⱦɫ��ѧ˵���Խ��͡�
�����Խ���������˵�ɽ�����ʵ�ֵ�Ⱦɫ���ؽ��dz���Ҫ�����Ǹ�����ĸ���������ϵ���Ч���ƣ���ͨ������������Ⱦɫ���ڵ���������ṩ�dz��ḻ�����Ŵ��ͣ�������ԶԶ������ͻ�����ṩ�ģ��Ա���Ȼѡ�����á�
����Ⱦɫ������һ���������ܴٽ����飨recombination�������ڳ�����ѵļ��������и�����ĸ��Ⱦɫ��Ķ����˶���1902����ǰ�ձ���Ϊ������ĸ����Ⱦɫ������Ϊ���Եĵ�����λ�˶�������ijЩѧ����Ϊ����ϸ���ij������ʱ���еĸ���Ⱦɫ�嶼�Լ������ʽ���ų���Ȼ��ͨ���ܾ������Ը�������Ⱦɫ������档�������������������ֳ���ڳ���ʱ�Ͳ���������壬Ȼ��������֤��������ֳ�Ѳ�������������Ҽ����γɵķ�ʽ��������ֳ�ѵ�û���κβ�ͬ�����⣬�ӺϵĴ���������и�����������ӡ����CarotherS��1913�������ھ��д�С��ͬ�ģ������ԣ�Ⱦɫ����������У��ϴ��Ⱦɫ�������������˶������Ǹ����ĺ�ĸ����Ⱦɫ���鲢����Ϊ��һ�ĵ�λ���з���ľ�����֤�ݡ�Ȼ����һ�ֺ������Ŵ���������������������mciotic drive������ֹȾɫ��������������������˵����ijЩ�������Ⱥ�б�����������������к����������
����Ⱦɫ������
����Ⱦɫ�����γɽ���ʱż������ȫ���ѣ�Ȼ���ٰ��µķ�ʽ������װ�����������ڶ��Ѵ�������һ��������������ζ��ѣ��м��һ�ο��Ե�ת�����������Ⱦɫ�塰��λ���������˿�����ڵ�λ��Ⱦɫ����Ͼ��DZ��ڵ�λ�������˿�����������Ⱦɫ����ھͳ�Ϊ�ۼ䵹λ����Ⱦɫ���ijһƬ�ζ������������ڣ�����룩��һ������ͬԴ��Ⱦɫ���Ͼͳ��֡���λ����������ʱҲ�����������ࣩ�Ƚ��������γ�������Ⱦɫ�壬����һ��Ⱦɫ�����ظ�������һ����ȱʧ�����������н�����˿���ģ�Ⱦɫ����ܲ��ϻ�һ���������м���˿�ϵģ�Ⱦɫ����ܽ��з��ѣ������ı仯��Ϊ��ѷ���š���Robertsonian rearrangements���������ԡ�ָ���Ƕ���Ⱦɫ���������Ŀ�������ϵ�Ⱦɫ�塣������ЩȾɫ��仯�ڽ����϶�����DZ�ڵ���Ҫ���壬��˿��Ҳû�н���Ⱦɫ���Ŵ�ѧ˵�ļ�ֵ�������Ŵ�Ӱ���Ⱦɫ������������Ϊ��Ⱦɫ��ͻ�䡱����������û����һ������ЩȾɫ��ͻ��ķ�����ʷ����Ϊ����˽�Ⱦɫ������������塣
����ijЩ��ʷѧ������Ħ�������о�С����Ⱦɫ���Ŵ�ѧ˵�Ĵ�ʼ������˵����Ȼ������ȷ��Ⱦɫ������Ե�֤ʵ������Ҫ�Dz������Ĺ��ף�����Ĺ���Ⱦɫ��ض����ɲ�ͬ���ʵ��Ŵ�����ֱ�����ж��ɵ��۵㡢�Լ��ϵ¶���������ķ��֣�����˵��Ȼ����1902��1904�굼��ɪ��-��������Ⱦɫ���Ŵ�ѧ˵����һѧ˵����������ͱ������ϸ��ѧ�ҽ��ܣ���Ϊ��ֻ��������ǰ20��ϸ��ѧ��չ�ı�Ȼ���
����������ѧ˵��˵������ǿȴ���ǿ�ҷ��ԣ������������������Ŵ�ѧ���籴��ɭ��Լ��ѷ���������Ħ�������ⲻ�ܲ�ʹ��ʷѧ�Ҹе��൱��㯡�����Ȼ�漰������ѧ��������Ҫѧ��֮�����ٹ̵ĸ�����硣����Ⱦɫ��ѧ˵�Ǽ�ӵظ������ͬ����ʵ���۶��ã�����������Ҫ�������֤�ݣ��ر���ʵ��֤�ݡ�����Ħ��С�鼰����ѧ����Ȼ�ṩ����Щ֤�ݣ�������1910���Ժ���£�������Ħ����Ⱦɫ��ѧ˵�ķ�����ת���֧���ߵ�ʱ��
����Ħ����1903�굽1910����һЩ����������������̵ع���Ⱦɫ��ѧ˵��Allen��1966�������ķ����۵�ܶ࣬����һ������ѧ˵ֻ�ǡ����ۡ���ȱ��ʵ����ݡ�Ħ����Ϊ����û�б�ʵ��֤ʵ�ľ��㲻�Ͽ�ѧ�����dz����ӡ�����������������Ҫ���ǣ������״���ɿ������ƶ�����Щ������λ�ڸ�����ͬ��Ⱦɫ�����Ǻ�������������ѧ˵�����£���ȫ��ͻ�ġ�
����Ȼ������1910��Ħ�������ڸ�ҹ֮��ת���Ⱦɫ��ѧ˵����Ҫ֧����֮һ���ṩ��һЩ���о����Ե�����֤�ݡ���������Ħ������һ����ת�䣿��һ����仯������ת��ǰ�����������µ����ڵõ�֤����
����1910��8�¡���������ѧ�ҡ���־������һƪĦ����1910a�����ҹ���Ⱦɫ��ѧ˵�ij���48ҳ�����ģ������յ�������2�£�����������Ħ�������������ۡ����ģ�1910b��������7��27�գ��յ�����7��23B��֮����������ڣ����һƪ����������Ħ����������Ⱦɫ��ѧ˵�ķ���̬�ȡ�
����1910��Ħ����48�꣬���Լ�ּ������ƣ��ͱ���ɭ�෴�����µ�ʵ�������ԭ�ȵĽ��Ͳ�����ʱ���ܹ��ı��Լ����뷨����һ���̶��ϣ�����Ȼ��Ħ����˼����Ȼ���ܵ�����Χ���������ǣ�������Ӱ�졣����һЩ����֤ʵ�˼���10��ǰ�������Ѻ�ͬ������ѷ��E��B��Wilson���������������������ѷ���۵��ֱ�Ħ����ɫ����������߶����������Щ�����˵���ɫ�����ʺ��Ը�Ķ�������û��Ħ����19����ƫִ��Ħ��С���Ա����Ҫ�ص�����Jack Schultz��1967��������Ҳ��Ħ��С��ĺ��ڳ�Ա֮һ����������ʵ�Ŀ̻�����Ħ���Ļ���̬�Ⱥ����յ�ϵͳ�۵㡭��˹���巶�ص�Խ���������Ͳ�����˹�ij�ɫʵ�鼼�ɡ������С������������Ա�����д�����˳��������С����Ӭ�ҡ��ڣ�����Ħ�����쵼�¹����������С����ĸ���Ա�о�����˭����Щ���ⷽ��ǿ�����ʵ��Ⱦɫ��ѧ˵���Ӳ鿼������Ҳ�ؽ�Ҫ���о�Ħ����ʷ��ר����˹���巶�أ�1965a����Allen��1967��1978�����о����յ���Carlson��1966��1974����Roll-Hansen��1978b����������Щ����Ӭ�ҵľ��Ķ����ԣ����Ƿdz���Ǣ����������䲢����Ϊһ֧����ʮ����������ü�˵һ�ݾ�����1911���Ժ�ܿ��������գ�������˹��˹���巶������ּ�˵����Ħ����һ������ؼ��Ҫ����Щ��˵����ͨ��ʵ�鳹�ؼ��Լ��顣
������ȻĦ�������������֣�����ȷ�ؽ��н��ͣ����������Լ�����ѧ˵����������֤�ݣ�Ȼ��Ҳ�кܶ���֤�ݱ�����������һ����ǿ�ġ�ת���ߡ�����ʱ��Ը����ص���ԭ����1910����ǰ��˼����ȥ��һֱ����1926�������������������ߵ�ƫ��ȴ�������о��Ŵ������ѧ�߹��ڻ����һ�н��۽����ԡ����ֵĺͶ��������ݡ�������ѧ˵������ȫ���������������������ʣ�����ѧ˵�������������ʶ��ԣ������ͺ�����Ⱦɫ���ϵ�λ�þ��ǻ����Ψһ���ʣ�
��������Ⱦɫ��ѧ˵�ķ�չ��Ѹ���ۻ����Ŵ�ѧ���ݵ�һ��������1915�����Ħ����˹���巶�أ����պͲ�����˹�����Ǻ����ġ��ϵ¶��Ŵ�ѧ�Ļ��ơ�һ���зdz���ȷ�ز����������Ϊʲô����ɭ��Լ��ѷ�Ȼ���������Ⱦɫ��ѧ˵�����˷ѽ⣬ΪʲôĦ�����������������֡�˹���巶�غͲ�����˹����û�к�����ѧ˵�����е���Ҫ�ø��µ�ʵ������֤Ⱦɫ��ѧ˵����ȷ�ԡ�����������̽�����ֱ����ϵ������ì��������Ա�֤����Щ������Ƕ�������Ⱦɫ��ѧ˵���Խ��͡��������е���֣�����Ϊʲô�����������������鱾ȥ�о������⡣���ҿ�������1915�굽1930���ڹ�Ӭ�Ŵ�ѧ�Ͼ��и߶ȴ�����һ˿�������о����������ϲ�û�ж�ɪ��-������ѧ˵����κ���Ҫ������������Щ�о�ȴ��֤����ѧ˵����ȷ�Բ�ָ������������ѧ�ϵ���Ҫ���塣
��������Ⱦɫ��ѧ˵Ϊʲô������˼��ҵķ������������о���ʱ���������ϾͿ����ҵ��𰸣�Coleman��1970��Roll-Han sen��1978b����Ⱦɫ��ѧ˵������������ѧ֪ʶ���õ�ֻʯƬ�ߣ�����Ҫ�ĵ������Ǽ�������ѧ�����ָ�����ͬ����ѧ�����ֶ���������۵�һ����֤�����ܾ����õ�ʵ���ϣ��Ӵ����ںϣ��Լ�19���͵�һЩ�������ۣ�����ϸ���˵���Դ����Ҳ��������������ѧ�ɵķ��磨����Coleman��1965��Churchill��1971������1910�굱ʱ���������˵��������������Ӫ����������һ��ӡ��һ��������������һ��������һ��̥ѧ����Ӫ����һ���ǿ������ߡ��ȳ����ߡ�ϸ��ѧ����Ӫ������������ʱ���õijƺ���1910��ʱ��������ܲ������С�������1800���Ժ���κ�һ���������ȳ����ߵı�ǩ��ʮ������������⡣����������ԭ�����Ǽ��˵Ļ�ԭ���ߣ������������Ƿ������ij̶Ȼ������������ߡ�����������Ҳ�ǻ�е���ߣ����������ิ��ˡ��������������������˶�����������ƫ�á���̬�����ͣ�������ͼ��һ�У���������������ֵ��ʾ�����������������ϲ�ͬ�Ŀ�������������ѧ�����ṹ����̬�������ԡ���ʷ�仯�Լ���Ⱥ���������͡����ǵġ������ԡ����ʹ�ʹ���������ڷ��ӣ�����ǻ�ѧ�������������Ӷ���������ѧ����
���������������ƺ���������Ӫ��ȷ����һ���Ͽ��������飬���������ڽ���������������������ϸ�����ͬ��һ��������һ�����ɵġ�����ɭ��Լ��ѷ�Լ�Ħ������������������ߣ����Ⱦɫ���Ŵ�ѧ˵����ȷ�ģ������ζ��Ҫ�������Լ��ĸ���ṹ���ҽ���ͼ˵�����������ϻ����������϶�����ˡ�
�������������߶Ա�����ϻ���ܿ������ԣ�����۵�е����ﲻ������������˵���������ȳ������ִ���縴��ȳ����������֮�������������Գ����ˣ�homunculus���������visviva������ʽ������Ȼ���ѽ�������Ȼ����̥ѧ������1816��1828�����ң�֮���ȳ��۵ij����˹۵����ڹ��ڻ����Ѳ��ٱ��˿��ǣ������Դ�����ѧ����ʶ���Ŵ�����ľ�ȷ�Ժ����۵�һ�㻯����������Ҳͬ��վ��ס�š����㣬κ˹���Ͳ�������˵���Ŵ��ľ�ȷ����ȻҪ��������ʵĽṹ�����Ŵ������ڽṹ�ϵĸ����ԣ���������Ϊɪ��-������Ⱦɫ��ѧ˵��������������������������Ƿ��ص�Bonnet�������ȳ�����ô�������������Ĺ۵㡣
�����������һ���Ƚϳ�ֵķ�������������̥ѧ��1883����������Ŵ����ʵ�������ѧ˵�ӱ����Ͽ����ܿ�ͱ��㱾�˵���Ƕ�����۵��Լ�ϸ����ϵ�о��Ľ������1890���һ����һ������̥ѧ�����ƺ������ױ�κ˹�������ʲ��ȷ���ѧ˵������������ϵ¶��ľ��ȷ���ȴ���ڽ��͡�����������ɪ�١�������ѧ˵֮��ı���ì�ܾ�����ʮ��ķ����о�������²��������
���������һ������������Դ�ڵ�һ�������Ŵ�ѧ˵���ڼ���1900������ڶ��Ŵ��ͺͱ�����֮����������������Ȼ�ȳ��۵ij�����ѧ˵�ѱ���������ijЩ��̥ѧ�Һ��Ŵ�ѧ�ҵ�˼������ȴ����һ��ģʽ���棬�������ÿһ����״���������е�ijһ���ض��Ŵ����Ӵ����ġ��Ŵ��Ϳ���˵�����͵ı����ͣ���Ȼ���dz�����ȴ���Ŵ�������������ѿ�����ӻ�����������Ƕ��ÿ����Ƕ����ڱ����͵��ض����֡������뷨�����������ϵ¶������ߵġ���λ��״�������С��¸��1889���ر�ָ�����Ӵ�ϸ��������ϸ���ʣ�����ϸ�����з��ӷ������á�����������ɷ�����ķ�����ɡ���������������˵��������Ŵ��������̬ѧ˵������ԭ���Ϻ��ϵij����˸��û��ʲô��ͬ������ɭ��Լ��ѷ���������Լ��Ĺ۵��ص�������Ⱦɫ��ѧ˵����̬ѧ���͡�
��������ʹκ˹��������ά���Լ��¹���̥ѧ������Ĵ����뷢��֮��Ĺ�ϵҲ�������á�Ħ������С�����������������ֿ������о����Ӵ����Ŵ�ѧ���֡�����ɭ����������ϸ��ѧ˵��ѧ����̳���κ˹����ͳ����Ҫ��һ��ͬʱ�ܹ����ʹ��ݺͷ������Ŵ�ѧ˵����������ָ�������֯�������к�����ȫ��ͬ��Ⱦɫ�壨����ֱ�����еĿ����Ի������ǿ��������۲쵽�ķ������������
����ֻҪ�������Ŵ��ͺͱ����ͣ��������߾Ͳ��ò����Ŵ����Ӻ�������״֮��һ��һ�Ĺ�ϵ��ij���ȳ��ۿ������⡣ijЩ���ϵ�λ��״ѧ˵��ѧ����Ϊij�������ж�����״���ж����Ŵ����ӡ������һ���Ժ��������Ƶ�κ˹��������һ�з������в�ͬ����״��Ȼ�в�ͬ�Ķ��ӣ����粻���dzɵ�����ϵĿ��Զ����仯��ÿһ��״����ͬ������ë���ÿ����״�����䶨�ӡ����ڻ����ٵ���Ϊ�Ŵ�����ͨ����ֳ������ֱ��ת��ɱ�������������Ȼ���£������ⲻ����һ�������۶��ҿ���˵��һ�ֱ�Ȼ�Ľ��ۡ���˵����������ֱ����ͷ����仯ʱ������֪�������������λ�������ã�������ֻ�ܰ���һ������һ����״��˵��һ�µĹ۵������ͣ�����ʹ������ˡ���Ⱦѧ˵������ǰ����
���������Ч�ԣ�Pleiotropy���Ͷ����Polygeny���ķ��֣������ģ����յ����ˡ���λ��״��ѧ˵�����������DZ���������������ʹ��Ⱦɫ��ѧ˵�����߰����˴����ȳ���Ӱ��Ӷ���������С������Ӫ֮��ĺ蹵��Ȼ���ⳡ��ս�����������Կ�����������ٵ�ȡ��ʤ�����������������ߵ�ѧ˵���Ϊ�Ŵ��ķ���ѧ˵��Carlson��1971�������Ϊ�����ڸ�������һλ�����Ŵ�ѧ�ң��Ȼ����ȷ�ģ������ղ����ǵ�һ�ˡ���������ǰ��κ˹�����¸��������������1880����ͺ����������ᵽ�Ŵ�����ķ��ӻ�����
��������ǿ��������ֻ�Ƕ�������Ӫ�����ۺ������ļ�����ԵĽ��ܡ�ÿ�������ߣ����籴��ɭ��Լ��ѷ������ά�˺�Ħ���������Լ��������Ϲ۵㣬ʵ������ʱ���൱�������ġ��˴�ì�ܵĻ�Ϲ۵㡣Ȼ��Ⱦɫ��ѧ˵���������ǵ����������һ�£����߲�һ�¡������һ�£����Ǿͱ�����߷��Ի��߷����������ӵ��������ɭ��Լ��ѷ����������̵Ŀ�ѧ�ҡ�
����Ⱦɫ���о�
�����ڲ�������E��B������ѷ�Ժ�Ⱦɫ���о���Ȼ�dz����гɹ���ϸ���Ŵ�ѧ����Ⱦɫ���о��ķ��ֺ��Ŵ�ѧ�о��ķ������ߵļ����ۺϣ��������й�����Ѹ�ٷ�չ��������пˣ�1929�������״�����Ⱦɫ��ķ����о���Heitz��Bauer��1933�����·���˫��Ŀ����ľ����Ⱦɫ�壬C��D��Darlinston���Ŵ�ϵͳ���о���M��J��D��White���о����Լ����������ϸ��ѧ�߶�����о���1970���Ⱦɫ���о��ֽ�����һ���µĻ�Ծʱ�ڡ�
������һ�������Ҫ��չ�Dz����˸����¼����Ľ��������Ŀǰͨ����֯������ʹϸ�������������ڵ�����Һ��ͬ��ʹϸ��������ˮ���ش��������ƷĴ����γɺ�ʹȾɫ���������ȼ�������Ⱦɫ����ĿԶ�ȹ�ȥ��ѹƬ����ȷ�ɿ���������ͨ���¼���ʹ����Ⱦɫ����Ŀ��48������Ϊ46�����ںܶ��о��У������������Ŵ����йػ���Ķ�λ����ȷ�������Ⱦɫ�嶼�dz���Ҫ��Ⱦɫ��������Ϻܸ��ӣ�ijЩ��ѧ���������в�ͬ��ֵ�Ӱ�������������Ⱦɫ���ϳ��ֲ�ͬ�Ĵ������������õ��Դ�������ͬ���Էֱ�Q����G����R����T����C��������Caspersson andZech 1972�����÷��������ʣ�밣�������֯��Ⱦɫ����Ա�ǿ��Եõ���һ����Ҫ��Ϣ��
������Щ�о�������Ҫ�ķ��ֿ������˽ԭ�����ϸ���������壩�ߵ����������ͬ���Ŵ����ʣ����ᣩ�������ɺ�����ɵ�Ⱦɫ������ͺߵ�����IJ�һ����Ȼ��������Ϊ��Щԭ�������DNA����RNA������֯�ṹ�dz��������ر��ʺ��ڽ���ijЩ���͵��Ŵ�ѧ�����������ǻ����ܺͻ�����ڿ��ơ����һֱ��1970�����ڷ����Ŵ�ѧ�Ĵ��о����Dz���ԭ��������ʵ����ϡ�
������ȻĿǰ������ԭ�������DNA��֯�ṹ���˽϶���˽⣬������������Ⱦɫ�廹�ܲ���������Cold Sprins HarborSymposia 1978����Ŀǰ��ֻ֪��DNA�����ڣ����룿�������ʣ��ر����鵰�ף������ϣ������м��������Щ�������ڻ�������Ͼ��о��������á�Ȼ�����ܽ������Ѿ������˴����й���ʵ�����ҿ������������Ⱦɫ����Ϊһ�����壬����Ҫ���һ���������Ľṹ���ܵ�����ѧ˵��Ϊʱ���磬���кܶ��Ҫ������ˣ������Ŵ������Ⱦɫ��ѧ˵������Ⱦɫ���о��Ľ�����������˵�Ǹոտ���Ⱦɫ���о���һ����ʱ����
����------------------
��������ѧԷ �ռ�����
ǰҳ
Ŀ¼
��ҳ